|
Yersinia pestis
Yersinia pestis, auch Pestbakterium oder Pestbazillus genannt, ist ein gramnegatives, unbegeißeltes, sporenloses, fakultativ anaerobes Stäbchenbakterium. Es zählt zu den Enterobakterien und ist der Erreger der Lungen- und Beulenpest. Yersinia pestis wurde von Alexandre Émile Jean Yersin 1894 entdeckt und von diesem Pasteurella pestis[1] genannt. Seit 1944 wird das bis dahin der Gattung Pasteurella zugerechnete Bakterium in die nach Yersin benannte Gattung Yersinia gestellt.[2] Vorgeschichte und GeschichteDie älteste bekannte DNA eines Stammes von Y. pestis wurde aus den Zähnen eines 5000 Jahre alten Jäger-und-Sammler-Skeletts in Lettland gewonnen. Der Tote war sorgsam beerdigt worden, ebenso wie drei andere Personen der gleichen Stätte, deren Zähne allerdings keine DNA-Spuren von Y. pestis aufwiesen. Hieraus schließen die Archäologen, dass der damalige Y.-pestis-Stamm nicht so hoch ansteckend und innerhalb von Tagen tödlich war wie die Stämme, die später große Epidemien, wie die Justinianische Pest oder die Pandemie des Mittelalters („Schwarzer Tod“) auslösten. Sie vermuten ein Bakterienreservoir in einheimischen Bibern (Castor fiber), die Träger eines verwandten Bakteriums sind und deren Überreste im Umkreis archäologischer Stätten der Region in großer Zahl gefunden werden.[3] VirulenzDie Virulenz von Yersinia pestis entsteht durch Exotoxin-, Endotoxin- und Bakterienkapselbildung. 1980 stellten Dan Cavanaugh und James Williams fest, dass die Virulenz des Bakteriums temperaturabhängig ist. Die Körpertemperatur des Flohs liege bei 24 °C, die des Menschen bei 37 °C und die der Ratte 1,5 °C höher. Der Temperaturunterschied zwischen Floh und Ratte könne die Virulenz des Bakteriums bei der Übertragung um fast das 50-Fache steigern. Dies liege an der Fähigkeit des Bakteriums, bei höheren Temperaturen Schutzmechanismen gegen die Phagocytose, einen Bestandteil der menschlichen Immunabwehr, zu entwickeln. Bei Temperaturen, wie sie im Körper des Flohs vorherrschen, werde dieser Schutz nicht aktiviert, aber 3 Stunden nach Eingang in einen Körper von 37 °C sei der Schutzmechanismus gegen Leukozyten und kurz danach derjenige gegen die Monozyten ausgebildet.[4] Von den hunderten von bekannten Bakterienstämmen sind nur wenige virulent. Die 40–50 Stämme von Yersinia pestis haben bei Mäusen eine große Bandbreite der Dosis, bei der die Hälfte der Versuchstiere stirbt (LD50), von unter 3 bis 100 Millionen.[5] Bricht der Krankheitserreger in die Blutbahn ein, was in 50–90 % der unbehandelten Fälle geschieht, so entsteht die Pestsepsis, also eine Streuung in die Blutbahn. Auf diesem Wege können praktisch alle Organe befallen werden. Unter anderem kommt es zur Lungenpest. Die Gefahr bei der Vermehrung innerhalb des Menschen ergibt sich aus der Ausbildung diverser Pathogenitätsfaktoren bei einer Umgebungstemperatur von 37 °C, also Körpertemperatur. So bildet Yersinia pestis bei dieser Temperatur eine antiphagozytär wirkende Kapsel aus, die als Fraktion 1 (F1) bezeichnet wird, und zwei weitere antiphagozytäre Antigene, die Virulenzantigene V und W. In Bezug auf die Virulenz des Erregers bedeutet dies, dass im Falle der Lungenpest die Übertragung von Mensch zu Mensch möglich ist, wenn es zu Kontakt mit hochinfektiösem Sputum eines an Lungenpest Erkrankten kommt. In diesem Fall kann sich innerhalb von Stunden eine primäre Lungenpest ausbilden. Systematik und AusbreitungR. Devignat teilte 1951 die Pestbakterien in 3 Hauptvarianten nach den starken biochemischen Unterschieden ein:
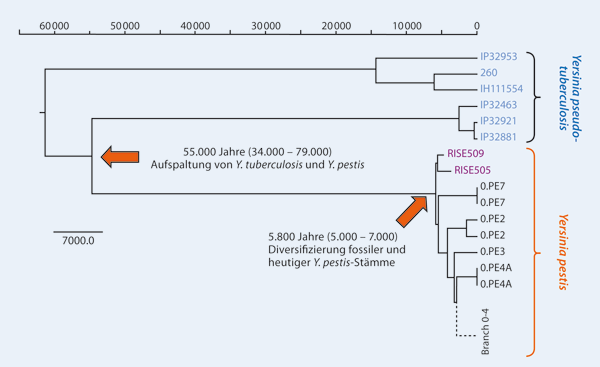
Dieses Modell war lange Zeit Grundlage für die Erklärung der Ausbreitungswege. Später stellte sich heraus, dass beide Hauptvarianten „Medievalis“ und „Antiqua“ in Kenia auftraten und „Orientalis“ und „Medievalis“ zusammen in der Türkei gefunden wurden.[7] Jeder dieser drei Pandemien wird einem Biovar zugeordnet, eine sich durch spezifische biochemische Eigenschaften unterscheidende Variante der Spezies Yersinia pestis. Mark Achtmann und andere kamen 1999 in ihren Studien zu dem Ergebnis, dass Yersinia pestis ein mutierter Klon seines nahen Verwandten Yersinia pseudotuberculosis ist, ein relativ harmloses Bakterium, das Magenbeschwerden verursachen kann und nur äußerst selten zum Tode führt. Die beiden Arten haben über 90 % des Genmaterials gemeinsam und würden nach taxonomischen Regeln als Varianten der gleichen Art bezeichnet, wenn sie nicht aus klinischen Gründen scharf unterschieden werden müssten. Die Autoren meinten, dass diese Variante höchstens 20.000 Jahre alt sein könne, das unsicherere Mindestalter aber wahrscheinlich bei 1000 Jahren liege.[8] 1997 wurde ein Pestbakterienstamm beschrieben, der multiresistent gegen Antibiotika war. Da er in älteren Proben nicht zu finden war, scheint er nach den Aussagen der Forscher erst 1995 entstanden zu sein.[9] Den ersten umfassenden durch Genomanalyse gestützten Blick auf etwa 1000 Isolate von Y. pestis gewannen Morelli und andere 2010 in einer Untersuchung, die zeigte, dass die ältesten pathogenen Stämme in China oder Russland entstanden sein mussten. Es fanden sich weitere Zusammenhänge in der Ausbreitung der Stämme mit dem Handel über die Seidenstraße, den Expeditionen von Zheng He und der dritten Pandemie von 1894.[10] 2011 schließlich hatten Bos, Schuenemann und andere die Gelegenheit, das Genom von Pestbakterien zu untersuchen, die aus Zähnen mittelalterlicher Londoner Leichen isoliert werden konnten. Ein Vergleich mit anderen bekannten Stämmen zeigte, dass es sich um Stämme handelte, die mit den ältesten Stämmen aus China am nächsten verwandt sein müssen. Das bedeutet beispielsweise, dass die mittelalterliche Epidemie tatsächlich von Asien ausging. Es bedeutet überraschenderweise auch, dass, wenn es wirklich noch frühere Pestepidemien gab, deren Stämme ausgestorben sind, da alle heute bekannten Stämme Abkömmlinge der Mittelalter-Variante sind.[11] Virulenzfaktoren und evolutionäre Veränderung des ErregersVerglichen mit seinem nächsten Verwandten, Yersinia pseudotuberculosis, hat Yersinia pestis im Laufe seiner Evolution die Fähigkeit verloren, im Boden zu überdauern. Im Gegenzug hat das Stäbchenbakterium zahlreiche neue Eigenschaften erworben, so diverse Virulenzfaktoren und Mechanismen zur Adaptation an entsprechende Säugetierwirte und parasitäre Vektoren.[12][13] Die Pest befiel bereits Gemeinschaften der späten Jungsteinzeit. Spuren des Erregers lassen sich in einem Zeitraum von vor 4800 bis etwa vor 3800 Jahren in Skeletten aus ganz Europa nachweisen.[14][15][16][3] Die frühen bronzezeitlichen Erreger konnten nur die primäre Lungenpest[A. 1] auslösen, weil ihnen das Gen ymt fehlte, welches für die stabile Besiedlung von Flöhen (Flohmagen) durch Y. pestis nötig ist.[16] So bildet der Pesterreger im Vektor Floh einen Biofilm, der wie ein Vormagenblock wirkt. Der Parasit kann damit nicht mehr abschlucken, wird hungrig und beißwütig und wechselt den Wirt. Dieser wissenschaftlich bestätigte Algorithmus bewirkt eine effiziente Verbreitung des Pesterregers auf andere Säugetierindividuen oder auch den Menschen. Die Genome von Y. pestis der frühen Epochen zeigten eine ausgestorbene Variante des Erregers, die auf das späte Neolithikum und die frühe Bronzezeit datiert werden konnte. Die Genome jener frühen Varianten sind in ihren genetischen Merkmalen mit den späteren Pesterregern nicht völlig identisch, so fehlen Virulenzfaktoren, die sich Vektoren (Flöhe) bedienen können und die ihre Verbreitung dadurch effizienter machen, durch die Fähigkeit, in Flöhen zu überleben, was den Hauptübertragungsweg der Krankheit auf Menschen und andere Säugetiere darstellt.[17] Im April 2022 wurden diese Ergebnisse bestätigt und gezeigt, dass es vor 2.500 in Eurasien zwei verschiedene Formen von Y. pestis gab.[18] Auch die im Mai 2023 veröffentlichten ca. 4.000 Jahre alten Funde in zwei englischen Gräbern (in Charterhouse Warren, Somerset und Levens, Cumbria) zeigten eine Variante von Y. pestis ohne ymt-Gen und ohne yapC-Gen. Letzteres verstärkt die Anheftung des Bakteriums an die Zellen und erleichtert vermutlich die Ausbreitung in der infizierten Person. Dazu passt, dass diese Personen offenbar nicht an der Pest, sondern an der Einwirkung äußerer Gewalt gestorben waren.[16] Damit sich die Pathogenität und Virulenz des Pesterregers über die natürlichen Nagetier-Reservoire und den Hauptvektor Floh ändern konnte, setzte es eine Veränderung (Mutation, Selektion) von Virulenzfaktoren voraus, die sowohl auf dem Bakterienchromosom als auch auf dessen Plasmiden kodiert sind.[19][20] Dabei unterschieden sich das Genom der frühen Peststämme von den späteren Varianten.[21][22] Wie bei allen Yersinien sind plasmidkodierte Virulenzfaktoren vorhanden. Zwei für Y. pestis spezifische Plasmide, „pMT1“ und „pPCP1“, tragen zahlreiche Virulenz-Gene, so bildet das Proteinprodukt eine Proteinkapsel aus, das sogenannte „F1-Antigen“. Dieses schützt Yersinia pestis vor der primären Immunantwort im Säugetierorganismus. Oder, eine membran-ständige Protease, ein „Plasminogen-Aktivator“, der die fibrinolytische Aktivität beeinflusst und dem Erreger über einen langen Zeitraum hinweg ermöglicht im intravasal zu persistieren und eine komplement-meditierte Zelllyse verhindert. Anderseits können andere Yersinia-assoziierte Faktoren zur Sepsis und unter Umständen zu einer Disseminierte intravasale Koagulopathie führen. Diese und weitere pathogene Eigenschaften verleihen Yersinia pestis seinen hochpathogenen Charakter. MutationenBei Yersinia sind es wohl 2 Plasmiden, die für pestis charakteristisch sind und seine Gefährlichkeit ausmachen. So wurde schon vermutet, dass eine plötzliche und umfassende Veränderung der Virulenz ein Hintergrundfaktor dafür gewesen sein kann, dass eine Pestepidemie plötzlich ausbrach und wieder verschwand, quer über die Jahrhunderte hindurch.[23] Gerade bei Viren und Bakterien ist eine Instabilität des Genmaterials zu beobachten, die zu vielen Mutationen führt. Inzwischen ist es gelungen, das gesamte Genom von Yersinia pestis zu entschlüsseln und zu kartografieren.[24] Dabei stellten die Forscher fest, dass das Bakterium genetische Besonderheiten aufweist, die auf häufige „intragenomische Rekombinationen“ hinweisen, dass es also in hohem Maße Gene anderer pathogener Organismen aufnehmen kann. Sie meinten, die Pestbakterien hätten Charakteristiken, die auf ständige Veränderungen hinwiesen.[24] Ein Jahr später entschlüsselte eine andere Forschergruppe einen anderen Bakterienstamm und bestätigte diese Einschätzung.[25] Annie Guiyoule und Bruno Rasoamanana untersuchten in Madagaskar die Gebiete mit besonders hoher Pestaktivität in den letzten Jahrzehnten. Sie isolierten 187 verschiedene Stämme aus der Zeit von 1926 bis 1996.[26] LebensdauerBei moderaten Temperaturen überlebt Y. pestis an den Mundwerkzeugen der Flöhe rund 3 Stunden.[27] Bereits 1944 ist festgestellt worden, dass Pestbakterien in Salzlösung und einer Temperatur um 25 °C bis zu 2 Wochen, bei 2–4 °C sogar bis zu zwei Jahre außerhalb eines Wirtstieres überleben und virulent sein können.[28] Experimente in der 1. Hälfte des 20. Jh. zeigten auch, dass Sonnenlicht die Bakterien rasch abtötet, die Lebensdauer jedoch stark von der Umgebung der Bakterien abhängt, insbesondere von deren Unterlage. In dünner Lauge auf Glas sterben die Bakterien innerhalb 1 Stunde ab, bei einer dicken Bakterienschicht vervierfachte sich die Lebensdauer und auf Stoff aus Hanf lebten sie bis zu 14 Stunden.[29] Während Temperaturen bei 55 °C tödlich sind, schaden ihnen niedrige Temperaturen nicht. In der Mandschurei wurden sogar noch virulente Bakterien in gefrorenen Pestleichen gefunden. Auch konnten die Bakterien fast einen Monat in Kornstaub überleben, der von infektiösen Ausscheidungen verunreinigt war.[30] Pestbakterien können Menschen bei Raumtemperatur und normaler Luftfeuchtigkeit in einem Zeitraum von mehr als 5 Tagen infizieren.[31] ÜbertragungFlöhe, insbesondere aber der Rattenfloh Xenopsylla cheopis können den Pesterreger übertragen. Aber auch Nosopsyllus fasciatus und der Menschenfloh Pulex irritans werden diskutiert, da Xenopsylla cheopis auf tropische Temperaturen angewiesen ist und in Europa nicht überleben kann. Flöhe sind blutsaugende Parasiten, die ihren Wirt direkt mit Y. pestis infizieren können. Wechselt der Rattenfloh von einem infizierten Nager – beispielsweise der Wanderratte oder der Hausratte – nach dessen Tod auf einen anderen Wirt, etwa Haustiere oder Menschen, ist er in der Lage, diese mit dem Pestbakterium zu infizieren. Dabei kann die Pesterkrankung für den Menschen ebenso tödlich sein wie für die Ratten. KrankheitserscheinungenAn der Bissstelle kommt es zu einer lokalen Infektion, die durch die Ausbildung einer Pustel gekennzeichnet ist und zur sogenannten Bubonenpest (Beulenpest) führt. Bei der septischen (blutvergiftenden) Form wird nicht immer eine Pestbeule entwickelt. Der Patient stirbt schnell ohne besondere äußere Symptome, aber mit einer hohen Bakterienkonzentration im Blut. Wenn diese Pestbakterien die Lunge angreifen, so wird diese Form als „sekundäre Lungenpest“ bezeichnet, die die gefährlichste Form mit dem raschesten Verlauf bildet. Neben hohem Fieber ist der blutige Auswurf eines der wenigen äußeren Symptome. Die blutvergiftende Wirkung wird ausgelöst, wenn die Bakterien ihren normalen Lebenszyklus vollenden und absterben. Dabei werden große Mengen toxischen Sekrets direkt in den Blutkreislauf abgegeben; Nieren und Leber können nekrotisch werden, wenn sie versuchen, das System von Toxinen zu reinigen. Am Ende erliegt das Opfer einem toxischen Schock. MeldepflichtIn Deutschland ist der direkte oder indirekte Nachweis von Y. pestis namentlich meldepflichtig nach § 7 des Infektionsschutzgesetzes (IfSG), soweit der Nachweis auf eine akute Infektion hinweist. Diese Meldepflicht für den Erreger betrifft in erster Linie die Leitungen von Laboren (§ 8 IfSG). In der Schweiz ist der positive und negative laboranalytische Befund zu Y. pestis für Laboratorien meldepflichtig und zwar nach dem Epidemiengesetz (EpG) in Verbindung mit der Epidemienverordnung und Anhang 3 der Verordnung des EDI über die Meldung von Beobachtungen übertragbarer Krankheiten des Menschen. Literatur
WeblinksCommons: Yersinia pestis – Album mit Bildern, Videos und Audiodateien
Anmerkungen
Einzelnachweise
|

{kind=link}