|
Biota del periodo Ediacárico Los organismos ediacáricos, también conocidos como biota del periodo Ediacárico o, anteriormente, biota o fauna vendiense, son antiguas formas de vida sésiles con forma tubular y de hoja que habitaron la Tierra durante el periodo Ediacárico (hace c. 635-542 millones de años) y que representan los organismos multicelulares complejos más antiguos conocidos dentro del nivel macrocelular de complejidad biológica.[nota 1] Aparecieron poco después de que concluyese la extensa y última glaciación del periodo Criogénico, y se extinguieron en su mayoría poco antes del abrupto incremento en la biodiversidad terrestre conocido como explosión cámbrica, que consistió en la primera aparición en el registro fósil de los patrones y morfologías básicas que más adelante supondrían la base de los animales modernos. Pocos elementos propios de la diversidad de los organismos ediacáricos fueron incorporados a estos patrones, y los distintos organismos del Cámbrico desplazaron y reemplazaron totalmente a las formas de vida que habían dominado el registro fósil Ediacara. Los organismos ediacáricos aparecieron por primera vez hace unos 580 millones de años y prosperaron a comienzos del Cámbrico, hace 542 millones de años. Aunque han sido descubiertos algunos fósiles que podrían representar supervivientes en estratos correspondientes al Cámbrico medio (hace c. 510-500 millones de años), las comunidades de fósiles más primitivas desaparecen del registro fósil al final del periodo Ediacárico, dejando únicamente fragmentos de los ecosistemas que anteriormente habían prosperado.[1] Existen diversas hipótesis para explicar esta desaparición, incluyendo un sesgo de conservación, un ambiente cambiante, la aparición de predadores o la competición con otras formas de vida. La posición filogenética de los organismos de esta biofacies constituye un gran problema para los paleontólogos. Mientras que muchos de ellos parecen corresponder a formas de vida sin representantes en la fauna actual, algunos organismos ediacáricos parecen haber estado estrechamente relacionados con grupos que más adelante se convertirían en prominentes; por ejemplo, la morfología de algunos taxones como Funisia dorothea sugieren su relación con los poríferos o cnidarios,[2] mientras que Kimberella muestra parecido con los moluscos e incluso ha sido propuesta como un antepasado de estos, lo que ha llevado a pensar que algunos organismos ya presentaban simetría bilateral, aunque esta afirmación es controvertida. La mayoría de los fósiles macroscópicos son morfológicamente distintos a las formas de vida posteriores: se asemejan a discos, bolsas llenas de barro o colchones esponjosos. Debido a las dificultades para situar en el árbol filogénico a estos organismos, algunos paleontólogos sugirieron al comienzo que al menos parte de la biota ediacárica representaría una rama independiente completamente extinta dentro de un reino propio, Vendobionta (inicialmente denominado como Vendozoa, nombre que implicaba una afinidad animal).[3] Otros, incluso, consideran que estos organismos formarían parte de una especie de «experimento fallido» de la vida multicelular, que supuso que la vida multicelular posterior volviera a evolucionar más adelante a partir de otros organismos unicelulares no relacionados.[4] La biota del periodo Ediacárico en su contexto
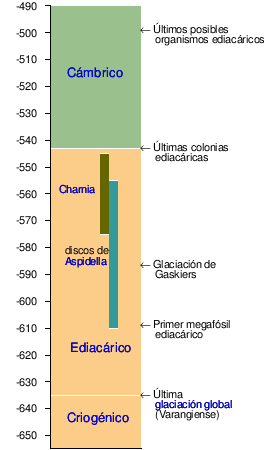
Escala del eje: hace millones de años.
HistoriaLos primeros fósiles ediacáricos descubiertos fueron los de Aspidella terranovica, con forma de disco, en 1868. Su descubridor, Alexander Murray, un agrimensor y geólogo, los consideró herramientas útiles para determinar la edad de rocas de la zona de la isla de Terranova.[7] Sin embargo, como se encontraban por debajo de los «estratos primordiales», los estratos cámbricos que se consideraba contenían los primeros indicios de vida, pasaron cuatro años antes de que alguien se atreviera a sugerir que podían ser fósiles. La propuesta de Elkanah Billings al respecto fue descartada por sus colegas a causa de su explicación sencilla, y los fósiles fueron considerados estructuras debidas al escape de gases, concreciones inorgánicas o incluso trampas colocadas por un dios malicioso para sembrar la falta de fe.[7] En aquellos tiempos todavía no se conocían estructuras similares en ningún otro lugar del mundo, y este debate pronto cayó en el olvido.[8] En 1933, Georg Gürich descubrió ejemplares muy similares a los de Terranova en Namibia,[9] pero la firme creencia en que la vida tenía sus orígenes en el Cámbrico hizo que fueran asignados a este periodo, y no se trazó ninguna relación con Aspidella. En 1946, Reg Sprigg descubrió fósiles similares a «medusa» en los montes Ediacara, pertenecientes a los montes Flinders de Australia,[10] pero se creía que estas rocas se habían formado en el Cámbrico inferior, de manera que, aunque el descubrimiento despertó cierto interés, no atrajo la atención de los especialistas.  No fue hasta el descubrimiento británico del icónico Charnia en 1957 cuando se empezó a considerar seriamente que el supereón Precámbrico hubiera albergado vida pluricelular diferenciada.[11] Este fósil en forma de fronda fue descubierto en el bosque de Charnwood, en Inglaterra,[12][13] y a causa de los detallados mapas geológicos del British Geological Survey no cupo ninguna duda que estos fósiles estaban insertos en rocas precámbricas. El paleontólogo Martin Glaessner trazó finalmente la conexión entre este descubrimiento y los anteriores,[14][15] y con una combinación de dataciones más precisas de los ejemplares existentes y una inyección de vigor a la investigación, se descubrieron muchos más miembros de la fauna ediacárica.[16] Sin embargo, todos los ejemplares descubiertos hasta 1967 se encontraban en yacimientos de arenisca muy rugosa que impide queden conservados los detalles delicados, haciendo que fuera difícil interpretarlos. El descubrimiento de S. B. Misra de capas de ceniza fosilíferas en las cenizas volcánicas de Mistaken Point (Terranova y Labrador) cambió la investigación, pues la conservación de los detalles que hacía posible la fina ceniza permitió describir características anteriormente inadvertidas.[17][18] La pobre comunicación, junto con la dificultad de correlacionar formaciones globalmente distintas, hizo que surgiera una plétora de nombres diferentes para estos organismos. En 1960, el nombre francés «Ediacarien», por los montes Ediacara en el sur de Australia, que derivan su nombre de la palabra aborigen Idiyakra, «el agua está presente», se propuso frente a los términos rivales «Sinian» y «Vendian»,[19] para referirse a las rocas precambrianas terminales, nombres que también fueron dados a los organismos. Finalmente, los términos «Edicaran» y «Ediacarian» se aplicaron a la época o período geológico precámbrico y sus rocas correspondientes. En marzo de 2004, la Unión Internacional de Ciencias Geológicas puso fin a la inconsistencia, estableciendo formalmente el nombre del periodo terminal del Neoproterozoico en honor del topónimo australiano.[20] ConservaciónLa vasta mayoría del registro fósil se compone de la robusta materia esquelética de cadáveres degradados. Por eso, como los organismos ediacáricos tenían cuerpos blandos y carecían de esqueleto, su abundante conservación es sorprendente. Probablemente contribuyó la ausencia de criaturas excavadoras en estos sedimentos,[21] pues a partir de la evolución de estos organismos en el Cámbrico, las impresiones de cuerpos blandos aparecen generalmente bioturbadas antes de que se pudieran fosilizar. Estromatolitos  Los estromatolitos son estructuras sedimentarias bioconstruidas originadas por colonias de microorganismos formando «tapetes», que secretan fluidos pegajosos (mucílago), inducen la precipitación de carbonatos o unen las partículas sedimentarias de diversas maneras. Parece que migran hacia arriba cuando quedan cubiertas por una capa delgada de sedimentos, pero eso es una ilusión provocada por el crecimiento de la colonia; los individuos en sí no se mueven. Si queda depositada una capa de sedimentos demasiado gruesa antes de que puedan crecer o reproducirse a través de ella, algunas partes de la colonia mueren, dejando fósiles con una característica textura arrugada de «piel de elefante».[22] La mayoría de los estratos ediacáricos con textura superficial rugosa en «piel de elefante», característica de los estromatolitos, contienen fósiles, además la mayoría de los fósiles de este periodo se encuentran en superficies con estromatolitos. Aunque los estromatolitos estuvieron muy extendidos en el pasado remoto, la evolución de sus predadores naturales (los «organismos pacedores») en el Cámbrico hizo disminuir significativamente su número,[23] por lo que estas comunidades están actualmente limitadas a refugios inhóspitos, donde los predadores no pueden sobrevivir el tiempo suficiente como para comérselas. FosilizaciónLa conservación de estos fósiles considerada un enigma científico fascinante, pues al ser organismos de cuerpo blando, normalmente no fosilizarían. A diferencia de otras formas de vida de cuerpo blando posteriores (como las de los esquistos de Burgess o las calizas de Solnhofen), los organismos ediacáricos no estaban localizados en ambientes restringidos sujetos a condiciones locales inusuales, sino que eran un fenómeno global. Por tanto, los procesos que intervinieron en la fosilización debieron ser sistemáticos y estar presentes en todo el mundo. Hubo de existir algo muy diferente en el periodo Ediacárico que permitió que estas delicadas criaturas fueran conservadas. La hipótesis más extendida es que los fósiles fueron preservados gracias a que fueron rápidamente cubiertos por cenizas o arena, atrapándolos junto al barro o los tapetes de microbios en los que vivían.[24] Las capas de ceniza ofrecen más detalles fósiles y pueden fecharse con precisión con un margen de error de un millón de años o menos por medio de la datación radiométrica.[25] Sin embargo, es más habitual encontrar fósiles ediacáricos en capas de arena depositadas por tormentas o por corrientes oceánicos de alta energía que arrastraban las capas superficiales del fondo marino, conocidos como turbiditas.[24] Actualmente, los organismos de cuerpo blando casi nunca se fosilizan así, pero la presencia de extensas capas de microbios probablemente contribuyó a la preservación de los organismos ediacáricos, estabilizando las impresiones en el sedimento.[26] ¿Qué fue conservado?El ritmo de cementación del sustrato superyacente, en relación con el ritmo de descomposición del organismo, determina si se conserva la superficie superior o inferior del organismo. La mayoría de fósiles en forma de disco se descompusieron antes de que se cubriera el sustrato superyacente, y la ceniza o la arena llenaron el vacío, dejando un molde de la parte inferior del organismo.[25] En cambio, los fósiles de organismos esponjosos tienden a descomponerse después de la cementación del sedimento superyacente, por lo que se conserva la superficie superior. Su naturaleza más resistente se refleja en el hecho de que, en raras ocasiones, se encuentran fósiles de organismos esponjosos dentro de estratos acumulados por tormentas. La violenta sedimentación no las destruyó, como lo habría hecho con los discos, menos resistentes. Además, en algunos casos, la precipitación bacterial de minerales formó una «máscara mortuoria» similar a un molde del organismo.[7] Morfología
Los organismos ediacáricos exhibían una inmensa variedad de características morfológicas. Su tamaño varía desde los milímetros a los metros; su complejidad, de simples «manchas» a otras mucho más complicadas; su rigidez, de robusta y resistente a análoga a la de las medusas. Casi todas las formas de simetría están presentes.[24] Estos organismos se distinguían de fósiles anteriores en que presentaban una estructura multicelular organizada y diferenciada, así como medidas de muchos centímetros. La variedad morfológica de los organismos ediacáricos puede agruparse grosso modo en los siguientes taxones «de forma»: embriones, discos, bolsas, organismos esponjosos, ediacáricos «no ediacáricos» e icnofósiles. EmbrionesLos descubrimientos recientes de vida multicelular precámbrica documentan numerosos casos de fósiles con forma de embriones, especialmente en la formación de Doushantuo, en China. Algunos hallazgos[29] despertaron un gran interés mediático,[30] aunque algunos investigadores han afirmado que en realidad son estructuras no orgánicas formadas por la precipitación de minerales dentro de un hueco.[31] Otros «embriones» han sido interpretados como restos de bacterias reductoras de azufre gigantes similares a Thiomargarita,[32] una propuesta en principio muy discutida, pero que está ganando aceptación.[33][34] Microfósiles de hace 632,5 millones de años —solo tres millones de años después del fin de las glaciaciones del Período Criogénico— podrían representar «etapas de reposo» embriónicas en el ciclo vital de los animales más antiguos conocidos.[35] Una teoría alternativa es que estas estructuras representan etapas adultas de los animales de este periodo.[36] Mediante el análisis de muestras de estos especímenes por microscopía tomográfica de rayos X de fuente sincrotrón, se ha visto que no se trata de embriones ni de ninguna otra estructura asociada a los metazoos, sino que más bien se trata de acumulaciones de células semejantes a quistes producto de una división celular de tipo politómico, es decir, que proceden de una célula progenitora que aumenta mucho de tamaño, produciéndose posteriormente divisiones celulares rápidas sucesivas con poco o ningún crecimiento entre etapas. Esta forma de división es característica de algunos protozoos. Según los autores del estudio, esto lleva a pensar que los llamados "embriones" de Doushantuo serían en realidad representantes de un grupo de holozoos ya extinguido, que utilizaban estas estructuras como propágulos.[37] DiscosFósiles con forma circular, como Ediacaria, Cyclomedusa y Rugoconites, hicieron que los fósiles ediacáricos fueran identificados originalmente como cnidarios, un grupo que también incluye las medusas y los corales.[10] Exámenes posteriores han conducido a interpretaciones alternativas de los fósiles en forma de disco: actualmente, ninguno es considerado con certeza una medusa. Otras interpretaciones proponen cercanía con los rizoides, protistas[38] y anémonas marinas; los patrones presentes donde se encuentran dos ejemplares han hecho que muchos individuos aparentes sean identificados como colonias de microbios[39][40] y otros pueden representar marcas de rayado formadas por organismos dotados de pedúnculos girando alrededor de sus agujeros.[41] Debido a que solo el dorso inferior del organismo se ha fosilizado, no se conocen las suficientes características útiles del organismo para establecer una estructura adecuada. BolsasFósiles como Pteridinium, conservados dentro de capas sedimentarias, se asemejan a «bolsas llenas de barro». La comunidad científica no ha llegado a un consenso sobre como interpretarlos.[42] Organismos esponjososLos organismos considerados en la definición revisada de Vendobionta de A. Seilacher[3] comparten una apariencia «esponjosa», y son semejantes a un colchón inflable. En algunas ocasiones estas «esponjas» se rompieron o resquebrajaron antes de conservarse; estos ejemplares dañados ofrecen indicios valiosos para el proceso de reconstrucción. Por ejemplo, las tres (o más) frondas petaloideas de Swartpuntia germsi solo se pueden reconocer en ejemplares dañados, pues normalmente, las diversas hojas quedaban ocultas cuando los organismos eran aplastados durante la rápida sedimentación.[43] Estos organismos parecen conformar dos grupos, los organismos «rangeomorfos» fractales y los «erniettomorfos», más sencillos.[44] Incluyendo en estos grupos los fósiles de los célebres Charnia y Swartpuntia, este tipo es el más característico de los organismos ediacáricos, pero también el más difícil de situar dentro del árbol filogenético conocido. Al no disponer de boca, intestinos, órganos reproductivos ni conservarse ninguna prueba de su estructura interna, su existencia resulta muy extraña según los parámetros actuales. La hipótesis más aceptada es que podían succionar los nutrientes del agua a su alrededor por ósmosis.[45] Ediacáricos «no ediacáricos»Algunos organismos ediacáricos han conservado detalles más complejos, lo que ha permitido que sean interpretados como posibles formas primitivas de filos vivientes, excluyéndolos de algunas definiciones de los organismos ediacáricos. El fósil más antiguo de este tipo es el supuesto Vernanimalcula, bilateral, que según algunos especialistas es, en cambio, la parte interior de un saco de huevos o un acritarco.[31][46] Ejemplos más recientes, aceptados casi universalmente como bilaterales, incluyen Kimberella, parecido a un molusco,[47] Spriggina,[27] y Parvancorina, con forma de escudo,[48] cuyas relaciones están actualmente en debate.[49] Una serie de fósiles conocidos como pequeños fósiles con caparazón están también representados en el periodo Ediacárico, el más célebre de los cuales es Cloudina,[50] un pequeño fósil en forma de tubo que presenta marcas de depredación, sugiriendo que, aunque los predadores quizás no habrían sido muy comunes durante el Ediacárico, como mínimo existían. En el periodo Ediacárico ya existían representantes de taxones modernos, algunos de los cuales se pueden reconocer en la actualidad. Las esponjas, las algas rojas y verdes, los protistas y las bacterias son fácilmente reconocibles, y algunos de ellos ya existían miles de millones de años antes del periodo Ediacárico. También se han descrito posibles artrópodos.[51] IcnofósilesCon la excepción de algunos escolitos muy simples,[52][53] los únicos cubiles conocidos del periodo Ediacárico son horizontales, en la superficie o justo debajo. Estos cubiles implican que existían organismos móviles con cabezas, que probablemente habrían presentado una simetría bilateral. Este hecho los situaría dentro del clado de los animales bilaterales,[52] pero también podrían ser consecuencia del movimiento de organismos más simples que rodaban mientras se alimentaban por el fondo marino.[cita requerida] Unos supuestos «cubiles» de hace 1100 millones de años podrían haber sido construidos por animales que se alimentaban de la parte inferior de los «tapetes» de microbios, lo que les habría protegido de un océano con características químicas poco aptas para la vida;[54] sin embargo, su anchura desigual y puntas estrechas hacen que sea difícil defender un origen biológico,[55] por lo que incluso los primeros proponentes de esta hipótesis ya no defienden su autenticidad.[56] Los cubiles observados implican un comportamiento simple, por lo que no parece tener continuidad la evidencia de un comportamiento de alimentación complejo y eficiente de principios del Cámbrico. Algunos fósiles ediacáricos, especialmente los discos, fueron interpretados en un principio como icnofósiles, pero esta hipótesis no fue confirmada. Además de los cubiles, se han encontrado icnofósiles directamente asociados con un fósil edicarense. Yorgia y Dickinsonia aparecen a menudo al final de largas vías de icnofósiles que se corresponden con su forma[57] o con un mecanismo de alimentación ciliar, pero el método de formación de estos fósiles no relacionados, pero que aparecen juntos sigue siendo, en su mayor parte, un misterio.[58] El potencial molusco Kimberella está asociado con marcas de estrías, posiblemente formadas por una rádula.[cita requerida] Clasificación e interpretaciónLa clasificación de los organismos ediacáricos es difícil, y existen diversas teorías en cuanto a la posición que les corresponde en el árbol filogenético.  CnidariaComo los cnidarios son considerados los eumetazoos (animales multicelulares con tejidos) más primitivos, el primer intento de categorizar estos fósiles los definió como medusas y pennatuláceos.[59] Sin embargo, un estudio detallado de su ciclo de crecimiento ha descartado esta hipótesis.[60][61] El amanecer de la vida animalMartin Glaessner sugirió en su libro The dawn of animal life (El amanecer de la vida animal, 1984) que los organismos ediacáricos eran miembros de un grupo corona de los filos modernos, pero que no eran similares porque todavía no habían desarrollado los rasgos característicos que se utilizan en la clasificación moderna.[62] Adolf Seilacher respondió sugiriendo que el periodo Ediacárico fue el escenario en que los animales superaron a los protistas gigantes como forma de vida dominante.[63] En 1986, Mark McMenamin afirmó que los organismos ediacáricos no disponían de una fase embrionaria, por lo cual no podían ser animales. Según él, estos organismos desarrollaron un sistema nervioso y un cerebro de forma independiente, de manera que «el camino hacia la vida inteligente comenzó más de una vez en este planeta» aunque esta idea no ha recibido mucho apoyo.[38] Nuevo filoLa afirmación más célebre de Adolf Seilacher fue que los organismos ediacáricos representaban un grupo monofilético único y extinguido de formas relacionadas que descendían de un antepasado común (un clado), por lo que propuso la creación del reino Vendozoa,[64][65] nombradas así por el periodo Vendiano, terminología actualmente obsoleta. Más adelante, excluyó algunos fósiles, que fueron identificados como metazoos, y volvió a proponer el filo «Vendobionta». Seilacher describió los vendobiontas como cnidarios esponjosos sin cnidoblastos. Esta ausencia impide que se alimentaran como los cnidarios actuales, de manera que sugirió que podrían haber sobrevivido gracias a una simbiosis con organismos fotosintéticos o quimiotróficos.[66]  LíquenesLa hipótesis de Gregory Retallack de que los organismos ediacáricos eran líquenes[67] no ha conseguido demasiado apoyo entre la comunidad científica.[68][69] Retallack argumenta que los fósiles no están tan aplastados como ocurre con las medusas fosilizadas en situaciones similares, y su relieve se aproxima más al de la madera petrificada. Remarca que las paredes quitinosas de las colonias de líquenes ofrecerían una resistencia similar a la compactación, y afirma que el gran tamaño de los organismos —a veces, de más de un metro de diámetro, mucho mayores que cualquiera de los cubiles conservados— representa una evidencia en contra de su clasificación dentro de los animales. Otras interpretacionesCasi todos los filos posibles han sido utilizados en alguna ocasión para acomodar los organismos ediacáricos,[70] desde las algas,[71] hasta las protistas conocidos como foraminifera,[72] pasando por los hongos,[73] o colonias de bacterias o microbios,[39] así como hipotéticos grupos intermedios entre las plantas y los animales.[74] OrigenCuando aparecieron los organismos ediacáricos, hace 655 millones de años, ya habían pasado casi 4000 millones de años desde la formación de la Tierra. Aunque ha supuesto fósiles que fechan de hace 3460 millones de años,[75][76] la primera prueba inequívoca de vida fecha de hace 2700 millones de años,[77] y hace 1200 millones de años ya existían con toda certeza células con núcleo.[78] Por eso cabe preguntarse por qué hizo falta tanto tiempo para que aparecieran formas con un grado de complejidad como el de los organismos ediacáricos. Posiblemente no haya ninguna explicación especial y el lento proceso evolutivo simplemente requirió 4000 millones de años para acumular las adaptaciones necesarias. De hecho, parece que hay un lento aumento en el nivel máximo de complejidad a lo largo de este tiempo, con la evolución de formas de vida más y más complejas paulatinamente, con indicios de vida anterior semi-compleja como Nimbia, hallada en la formación Twitya y fechada hace 610 millones de años,[79] que posiblemente presentaba la morfología más compleja de aquel tiempo.  Una corriente de pensamiento alternativo es que simplemente no era una ventaja ser grande hasta la aparición de los organismos ediacáricos; el ambiente favorecía las formas pequeñas. Un ejemplo actual de este hecho es el plancton, cuyo pequeño tamaño le permite reproducirse rápidamente para aprovechar los abundantes, pero efímeros nutrientes de los florecimientos de algas. Sin embargo, el ambiente tendría que haber sido muy diferente a todos los conocidos para que una dimensión mayor no hubiera sido nunca ventajosa. Uno de los principales factores que limitan el tamaño es la cantidad de oxígeno de la atmósfera. Sin un sistema circulatorio complejo, las bajas concentraciones de oxígeno hacen muy difícil su asimilación por parte de un organismo lo bastante rápido como para responder a sus necesidades metabólicas. En la Tierra primitiva, los elementos reactivos como el hierro y el uranio existían solo en forma reducida y podían reaccionar, oxidándose, con el oxígeno producido por organismos fotosintéticos. El oxígeno no pudo empezar a acumularse en la atmósfera hasta que todo el hierro quedó oxidado (formando bandas de hierro), así como los demás elementos oxidables. Donald Canfield detectó indicios de las primeras cantidades significativas de oxígeno en la atmósfera justo antes de la aparición de los primeros fósiles ediacáricos,[80] por lo que la presencia de oxígeno atmosférico pronto fue señalada como el posible elemento desencadenante de evolución divergente del periodo Ediacárico.[81] Parece que el oxígeno se acumuló en dos etapas: la aparición de pequeños organismos sésiles parece corresponderse con un primer impulso de oxigenación, y la aparición de organismos mayores y móviles coincide con un segundo.[82] Sin embargo, la resolución del registro fósil conocido es demasiada baja para considerar esta teoría como definitiva, y las investigaciones actuales intentan determinar con precisión el papel que podría haber jugado el oxígeno, pues la anoxia tuvo un efecto muy limitado en otras etapas con vida multicelular, como durante el Cámbrico temprano o el Cretácico.[83] Los periodos de frío intenso también han sido sugeridos como un obstáculo para la evolución de vida multicelular. Los embriones más antiguos conocidos, procedentes de la formación de Doushantuo en China, aparecen solo un millón de años después de que la Tierra emergiera de una glaciación global, sugiriendo que la capa de hielo y los fríos océanos podrían haber impedido la aparición de vida compleja.[84] Es posible que hubiera evolucionado vida multicelular antes de estas glaciaciones, y que estas lo extinguieran. Sin embargo, la diversidad de formas de vida en la Antártida actual plantea desacuerdos razonables en cuanto a si las temperaturas frías aumentan o reducen el ritmo de la evolución. A comienzos del año 2008 un equipo analizó la disparidad de las estructuras de organismos ediacáricos a partir de tres fósiles diferentes: Avalon, en Terranova y Labrador, Canadá, fechado en 575 a 565 millones de años, mar Blanco, en Rusia, (560-550 millones de años) y Nama, en Namibia (550-542 millones de años), inmediatamente antes del comienzo del Cámbrico. Concluyeron que, mientras que las muestras del mar Blanco presentaban la mayor parte de las especies, no había diferencias significativas entre los tres grupos, por lo que antes del periodo de Avalon estos organismos debieron sufrir su propia «explosión», similar a la explosión del Cámbrico.[85] DesapariciónLa baja resolución del registro fósil hace que la desaparición de los organismos ediacáricos sea en parte un misterio. Parece que hubo una desaparición relativamente abrupta a finales del periodo Ediacárico. La presencia de organismos «ediacáricos» en el Cámbrico no tiene una aceptación universal. La causa —y realidad— de esta desaparición está actualmente en debate. Sesgo de conservaciónLa abrupta desaparición de fósiles ediacáricos cerca del límite del Cámbrico inferior podría deberse simplemente al hecho de que las condiciones ya no favorecían la fosilización de los organismos ediacáricos, que podrían haber continuado prosperando sin conservarse.[22] Sin embargo, si eran comunes, se tendría que haber encontrado más de un ejemplar ocasional[1] en conjuntos fosilíferos excepcionalmente bien preservados (yacimientos como los esquistos de Burgess o los esquistos de Maotianshan),[86] a menos que estos yacimientos representen un ambiente que los organismos ediacáricos nunca habitaron, o no apto para su conservación.  Depredación y organismos pacedoresSe ha sugerido que, en el Cámbrico inferior, organismos en una posición más alta dentro de la cadena trófica causaron que prácticamente desaparecieran las capas microbianas. Si estos organismos «pacedores» aparecieron en la misma época en la que empezaron a declinar los organismos ediacáricos, se puede sugerir que los «pacedores de bacterias» desestabilizaron el sustrato microbiano, causando como mínimo un desplazamiento de los organismos de la biota ediacárica o perturbando hasta tal punto el ecosistema, que condujera a su extinción. Otra posibilidad es que los nuevos animales con esqueleto podrían haberse alimentado directamente de los organismos ediacáricos, relativamente indefensos.[38] Sin embargo, si la interpretación de Kimberella (ediacárico) como «organismo pacedor» es correcta, se puede deducir que los organismos ediacáricos ya afrontaban una amenaza limitada de «depredación».[47] Sin embargo, hay poca evidencia de icnofósiles del periodo Ediacárico, hecho que a priori se opone a la teoría de los pacedores activos. Además, el inicio del Cámbrico se define por la aparición de un ensamblaje global de icnofósiles, bien diferente del propio del periodo Ediacárico, con pocos indicios de actividad.  CompetenciaEs posible que un aumento de la competencia a causa de la evolución de innovaciones clave dentro de otros grupos, quizás como respuesta a la depredación,[21] expulsase a los organismos ediacáricos de sus nichos ecológicos. Sin embargo, este argumento no ha explicado con éxito fenómenos similares. Por ejemplo, la «exclusión competitiva» de los braquiópodos por parte de los bivalvos fue finalmente considerada el resultado coincidente de dos tendencias no relacionadas.[87] Cambio en las condiciones ambientalesAunque es difícil inferir el efecto que tienen unas condiciones planetarias cambiantes sobre los organismos, comunidades y ecosistemas, hubo grandes cambios a finales del Precámbrico y principios del Cámbrico inferior. La fragmentación de los supercontinentes,[88] la subida del nivel del mar (creando mares someros y «aptos para la vida»),[89] una crisis de nutrientes disponibles,[90] fluctuaciones en la composición de la atmósfera, incluyendo los niveles de oxígeno y dióxido de carbono,[91] y cambios en la química de los océanos[92] que potenciasen la biomineralización[93] son factores que pudieron haber colaborado en la extinción de la biota ediacárica. Asociaciones ediacáricasSe conocen fósiles de tipo Ediacara en veinticinco localidades de todo el mundo y en una gran variedad de condiciones de deposición,[20] por lo que se los suele agrupar en tres tipos principales (asociaciones), que reciben el nombre de las localidades típicas. Cada asociación tiende a ocupar su propia región de morfoespacio y, después de un impulso inicial de diversificación, no cambia mucho durante el resto de su existencia.[94] Asociación de tipo AvalonLa asociación de tipo Avalon se define respecto al yacimiento de Mistaken Point, en Canadá, el primer lugar donde se identificó un gran número de fósiles ediacáricos.[95] Es fácil datar la asociación porque contiene muchas capas finas de ceniza, que son una buena fuente de zircón, utilizado en el método de datación radiométrica de uranio-plomo. Estas finas capas de ceniza también preservan detalles muy delicados. Los miembros de esta biota particular parecen haber sobrevivido hasta la extinción de todos los organismos ediacáricos al comienzo del Cámbrico.[94]
Los organismos incluyen rangeomorfos de las profundidades marinas, como Charnia, que comparten un ciclo de crecimiento fractal.[97] Es probable que fueran preservados in situ (es decir, no cambiaron de posición después de morir), aunque esta premisa no tiene una aceptación universal. Esta asociación, a pesar de ser menos diversa que la de tipo Ediacara o Nama, se parece a las comunidades carboníferas de alimentación en suspensión, lo que puede sugerir la presencia de alimentación por filtraje,[98] pues según la mayoría de las interpretaciones los organismos se encontraban en aguas demasiado profundas para permitir la fotosíntesis. La baja diversidad puede reflejar la profundidad del agua —que limitaría las posibilidades de especiación— o simplemente podría ser demasiado joven para la presencia de una gran biodiversidad. La opinión actual está dividida entre estas dos hipótesis.[96] Asociación de tipo EdiacaraLa asociación de tipo Ediacara debe su nombre a las colinas Ediacara de Australia, y consiste en fósiles preservados alrededor de zonas cerca de desembocaduras de ríos (haces prodeltaicos).[94] Se los suele encontrar en capas de arena y de limo formadas por debajo de la base habitual de moción acuática generada por las oleadas, pero en aguas lo bastante someras como para ser afectadas por las oleadas durante tormentas. La mayoría de fósiles están conservados como huellas en capas microbianas, pero algunos están preservados dentro de niveles de arena.[96] Asociación de tipo NamaLa asociación de tipo Nama se define según el yacimiento homónimo en Namibia. El tipo de fósiles más habitual consiste en una conservación tridimensional, con organismos preservados camas de arena con capas interiores. Dima Grazhdankin opina que estas formas representan organismos excavadores,[42] mientras que Guy Narbonne sostiene que vivían en la superficie.[99] Estas capas quedan en medio de unidades que comprenden areniscas, limos y esquistos, además de capas microbianas que en el caso de aparecer suelen contener fósiles. El ambiente es interpretado como barreras de arena formadas en la boca de los defluentes de un delta fluvial.[96] Significado de las asociacionesEn la región del mar Blanco de Rusia se han encontrado los tres tipos de asociación en lugares muy próximos. Este hecho, sumado al considerable solapamiento en el tiempo de los organismos, hace que sea improbable que representen estadios evolutivos o comunidades distintas en el tiempo. Como se encuentran por todo el mundo —en todos los continentes excepto la Antártida— los límites geográficos no parecen tener influencia;[100] se encuentran los mismos fósiles a todas las paleolatitudes (la latitud en que se formó el fósil, teniendo en cuenta la deriva continental) y en diferentes cuencas sedimentarias.[96] Lo más probable es que las tres asociaciones representen organismos adaptados para sobrevivir en ambientes diferentes, y que cualquier patrón aparente en diversidad o en edad no sea más que una impresión provocada por las pocas muestras descubiertas, pues los yacimientos fosilíferos ediacáricos son muy escasos. Como los organismos ediacáricos representan un estadio primitivo en la historia de la vida multicelular, no es sorprendente que no estén ocupados todos los diferentes modos de vida.[96] Se ha calculado que, de los noventa y dos modos de vida potencialmente posibles (combinaciones de modo de alimentación, posición en la cadena trófica, movilidad…), solo una docena estaban ocupados a finales del Ediacárico. En la asociación de tipo Avalon solo se encuentran cuatro.[101] La falta de depredación a gran escala y la excavación vertical son probablemente los factores más importantes que limitaban la diversidad ecológica; la aparición de esta durante el Cámbrico inferior permitió que el número de modos de vida ocupados aumentara a treinta. Véase tambiénNotasReferencias
Bibliografía
Enlaces externos
|
|||||||||||||||||||






{kind=link}
{kind=link}