| Name
|
Year
|
Formation
|
Location
|
Notes
|
Images
|
| Abdarainurus
|
2020
|
Alagteeg Formation (Late Cretaceous, Campanian)
|
 Mongolia Mongolia
|
Inconsistent in phylogenetic placement. Could represent an unknown lineage of macronarians[1]
|

|
| Abrosaurus
|
1989
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
 China China
|
Had unusually large fenestrae
|

|
| Achillobator
|
1999
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Its robust build suggests it was not a cursorial animal[2]
|

|
| Adasaurus
|
1983
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Its sickle claw was markedly reduced compared to other dromaeosaurids
|

|
| Aepyornithomimus
|
2017
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
The first ornithomimosaur named from a dry desert environment
|

|
| Agilisaurus
|
1990
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
The holotype specimen was discovered during the construction of the museum where it is now housed
|

|
| Albalophosaurus
|
2009
|
Kuwajima Formation (Early Cretaceous, Valanginian to Hauterivian?)
|
 Japan Japan
|
Only known from fragments of a skull
|

|
| Albinykus
|
2011
|
Javkhlant Formation (Late Cretaceous, Santonian)
|
Mongolia
|
Preserved in a sitting position not unlike that of modern birds
|

|
| Alectrosaurus
|
1933
|
Bayan Shireh Formation?, Iren Dabasu Formation (Late Cretaceous, Cenomanian to Santonian?)
|
China
Mongolia?
|
Had long legs which may be an adaptation to pursuit predation[3]
|

|
| Alioramus
|
1976
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Possessed an elongated snout with a row of short crests
|

|
| Almas
|
2017
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Preserved alongside eggshells which may have come from a troodontid[4]
|

|
| Alpkarakush
|
2024
|
Balabansai Formation (Middle Jurassic, Callovian)
|
 Kyrgyzstan Kyrgyzstan
|
Distinguishable by its uniquely apparent, rugose orbital brow[5]
|

|
| Altirhinus
|
1998
|
Khuren Dukh Formation (Early Cretaceous, Barremian to Albian)
|
Mongolia
|
Had a distinctively elevated nasal bone which supported a large nasal cavity
|

|
| Alxasaurus
|
1993
|
Bayin-Gobi Formation (Early Cretaceous, Albian)
|
China
|
Most of the skeleton is known, which allowed researchers to connect therizinosaurs to other theropods
|

|
| Ambopteryx
|
2019
|
Unnamed formation (Late Jurassic, Oxfordian)
|
China
|
Preserves stomach contents containing gastroliths and bone fragments, suggesting an omnivorous diet
|

|
| Amtocephale
|
2011
|
Bayan Shireh Formation (Late Cretaceous, Turonian to Santonian)
|
Mongolia
|
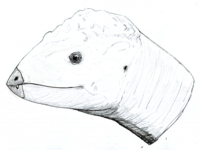
One of the oldest known pachycephalosaurs
|
|
| Amurosaurus
|
1991
|
Udurchukan Formation (Late Cretaceous, Maastrichtian)
|
 Russia Russia
|
One specimen may have come from an individual with a limp[6]
|

|
| Analong
|
2020
|
Chuanjie Formation (Middle Jurassic, Bajocian)
|
China
|
Originally described as a specimen of Chuanjiesaurus but it was assigned to a new genus due to several morphological differences
|

|
| Anchiornis
|
2009
|
Tiaojishan Formation (Late Jurassic, Oxfordian)
|
China
|
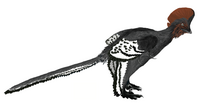
Analysis of fossilized melanosomes suggests a mostly gray or black body, white and black patterns on its wings and a red head crest[7]
|
|
| Anhuilong
|
2020
|
Hongqin Formation (Middle Jurassic, Aalenian to Callovian)
|
China
|
Closely related to Huangshanlong and Omeisaurus, all forming an exclusive clade of mamenchisaurids
|

|
| Anomalipes
|
2018
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
May have been closely related to Gigantoraptor despite its significantly smaller size[8]
|

|
| Anserimimus
|
1988
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Had powerful forelimbs with uniquely straight, flattened claws
|

|
| Aorun
|
2013
|
Shishugou Formation, (Late Jurassic, Oxfordian)
|
China
|
Potentially a basal member of the alvarezsaurian lineage[9]
|

|
| Aralosaurus
|
1968
|
Bostobe Formation, (Late Cretaceous, Santonian to Campanian)
|
 Kazakhstan Kazakhstan
|
Its crest has been interpreted as being arch-shaped as in kritosaurin hadrosaurids, but this cannot be confirmed
|

|
| Archaeoceratops
|
1997
|
Xinminbao Group (Early Cretaceous, Aptian)
|
China
|
Had no horns and only the beginnings of a frill
|

|
| Archaeocursor
|
2024
|
Ziliujing Formation (Early Jurassic, Sinemurian to Pliensbachian)
|
China
|
The oldest and most primitive ornithischian from Asia
|

|
| Archaeornithoides
|
1992
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Known from only a partial skull with scratches that may have been created by a small mammal[10]
|

|
| Archaeornithomimus
|
1972
|
Bissekty Formation?, Iren Dabasu Formation (Late Cretaceous, Cenomanian to Coniacian?)
|
China
 Uzbekistan? Uzbekistan?
|
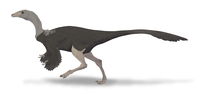
Unlike other ornithomimosaurs, its feet were not arctometatarsalian
|
|
| Arkharavia
|
2010
|
Udurchukan Formation (Late Cretaceous, Maastrichtian)
|
Russia
|
Described from a series of vertebrae, several of which were found to not belong to this taxon[11]
|

|
| Arstanosaurus
|
1982
|
Bostobe Formation (Late Cretaceous, Santonian to Campanian)
|
Kazakhstan
|
Poorly known
|

|
| Asiaceratops
|
1989
|
Khodzhakul Formation, Xinminbao Group? (Early Cretaceous? to Late Cretaceous, Aptian? to Cenomanian)
|
China?
Uzbekistan
|
Potentially a leptoceratopsid[12]
|

|
| Asiatosaurus
|
1924
|
Öösh Formation, Xinlong Formation (Early Cretaceous, Barremian to Albian)
|
China
Mongolia
|
Two species have been named but both are only known from extremely scant remains
|

|
| Asiatyrannus
|
2024
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Small for a tyrannosaurid although the holotype belongs to an immature individual[13]
|

|
| Auroraceratops
|
2005
|
Xinminbao Group (Early Cretaceous, Albian)
|
China
|
Known from more than eighty specimens, including complete skeletons
|

|
| Aurornis
|
2013
|
Tiaojishan Formation (Late Jurassic, Oxfordian)
|
China
|
If an avialan as originally described, it would be one of the oldest members of the group
|

|
| Avimimus
|
1981
|
Barun Goyot Formation, Nemegt Formation (Late Cretaceous, Campanian to Maastrichtian)
|
Mongolia
|
Bonebed remains indicate a gregarious lifestyle. It may have formed age-segregated herds for lekking or flocking purposes[14]
|

|
| Bactrosaurus
|
1933
|
Iren Dabasu Formation, Majiacun Formation? (Late Cretaceous, Cenomanian to Santonian?)
|
China
|
Remains of at least six individuals are known, making up much of the skeleton
|

|
| Bagaceratops
|
1975
|
Barun Goyot Formation, Bayan Mandahu Formation, Djadochta Formation? (Late Cretaceous, Campanian)
|
China
Mongolia
|
May have been a direct descendant of Protoceratops which it physically resembles[15]
|

|
| Bagaraatan
|
1996
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
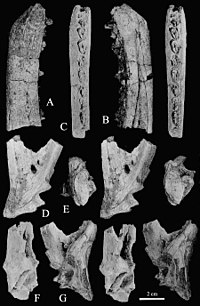
Recently found to be chimeric as some bones actually come from a caenagnathid[16]
|
|
| Bainoceratops
|
2003
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Its supposedly diagnostic features may fall within Protoceratops variation[17]
|

|
| Baiyinosaurus
|
2024
|
Wangjiashan Formation (Middle Jurassic, Bathonian)
|
China
|
Exhibits anatomical characteristics transitional between basal thyreophorans and derived stegosaurs
|

|
| Banji
|
2010
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Vertical striations adorned the sides of its crest
|

|
| Bannykus
|
2018
|
Bayin-Gobi Formation (Early Cretaceous, Barremian to Aptian)
|
China
|
Exhibited a transitional hand morphology for an alvarezsaur, having three fingers of roughly equal length with the first one being robust
|

|
| Baotianmansaurus
|
2009
|
Gaogou Formation (Late Cretaceous, Cenomanian to Turonian)
|
China
|
Large but known from only a few bones
|

|
| Barsboldia
|
1981
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Possessed elongated neural spines particularly above the hips
|

|
| Bashanosaurus
|
2022
|
Shaximiao Formation (Middle Jurassic, Bathonian)
|
China
|
Its skeleton combines traits of stegosaurs and more basal thyreophorans
|

|
| Bashunosaurus
|
2004
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
Although described as a macronarian, this has yet to be rigorously tested[18]
|

|
| Batyrosaurus
|
2012
|
Bostobe Formation (Late Cretaceous, Santonian to Campanian)
|
Kazakhstan
|
Remains originally identified as Arstanosaurus
|

|
| Bayannurosaurus
|
2018
|
Bayin-Gobi Formation (Early Cretaceous, Aptian)
|
China
|
Known from a well-preserved, almost complete skeleton
|

|
| Beg
|
2020
|
Ulaanoosh Formation (Early Cretaceous to Late Cretaceous, Albian to Cenomanian)
|
Mongolia
|
Its preserved skull has a rugose texture
|

|
| Beibeilong
|
2017
|
Gaogou Formation (Late Cretaceous, Cenomanian to Coniacian)
|
China
|
Similar to but more basal than Gigantoraptor.[19] Known from only a single embryo still in its egg
|

|
| Beipiaosaurus
|
1999
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Preserves evidence of downy feathers as well as a secondary coat of simpler "elongated broad filamentous feathers" or EBFFs[20]
|

|
| Beishanlong
|
2010
|
Xinminbao Group (Early Cretaceous, Aptian)
|
China
|
Lacked the elongated claws of more derived ornithomimosaurs
|

|
| Bellusaurus
|
1990
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Known from a bone bed with the remains of seventeen juvenile specimens
|

|
| Bienosaurus
|
2001
|
Lufeng Formation (Early Jurassic, Sinemurian)
|
China
|
Potentially synonymous with Tatisaurus[21]
|

|
| Bissektipelta
|
2004
|
Bissekty Formation (Late Cretaceous, Turonian to Coniacian)
|
Uzbekistan
|
Analysis of its braincase suggests poor hearing and eyesight but good olfaction and taste. It has been suggested to be a filter feeder[22]
|

|
| Bolong
|
2010
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Originally known from only a skull. An almost complete skeleton was described in 2013[23]
|

|
| Borealosaurus
|
2004
|
Sunjiawan Formation (Late Cretaceous, Cenomanian to Turonian)
|
China
|
Its caudal vertebrae were distinctively opisthocoelous
|

|
| Borogovia
|
1987
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
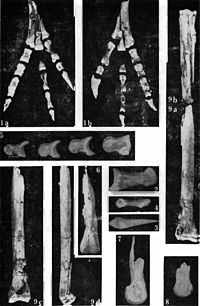
Had a uniquely straight, flattened sickle claw, which may have had a weight-bearing function
|
|
| Breviceratops
|
1990
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Only known from juvenile remains but can be distinguished from other protoceratopsids
|

|
| Brohisaurus
|
2003
|
Sembar Formation (Late Jurassic, Kimmeridgian)
|
 Pakistan Pakistan
|
Possibly an early titanosauriform
|

|
| Byronosaurus
|
2000
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Two juvenile skulls were found in an oviraptorid nest and claimed to be evidence of nest parasitism in this taxon, but both their identity and taphonomy have been questioned[4][24]
|

|
| Caenagnathasia
|
1994
|
Bissekty Formation (Late Cretaceous, Turonian to Coniacian)
|
Uzbekistan
|
One of the oldest and smallest known caenagnathoids
|

|
| Caihong
|
2018
|
Tiaojishan Formation (Late Jurassic, Oxfordian)
|
China
|
Possessed platelet-shaped melanosomes that produced iridescence as in modern trumpeters
|

|
| Caudipteryx
|
1998
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Two species are known. At least C. zoui did not have secondary feathers attached to the lower arm
|

|
| Ceratonykus
|
2009
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Several osteological features were described as similar to ornithischians[25]
|

|
| Changchunsaurus
|
2005
|
Quantou Formation (Early Cretaceous to Late Cretaceous, Aptian to Cenomanian)
|
China
|
Had wavy enamel on its leaf-shaped teeth that made them more resistant to wear. This feature is also present in hadrosaurs[26]
|

|
| Changmiania
|
2020
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Preserved in a curled up position as if it was sleeping in a potential burrow
|

|
| Changyuraptor
|
2014
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
The largest microraptorian dromaeosaurid known. Had tail feathers almost a foot long[27]
|

|
| Chaoyangsaurus
|
1999
|
Tuchengzi Formation (Late Jurassic, Tithonian)
|
China
|
Known by a number of alternate spellings (e.g. Chaoyangosaurus, Chaoyoungosaurus) before its formal description
|

|
| Charonosaurus
|
2000
|
Yuliangze Formation (Late Cretaceous, Maastrichtian)
|
China
|
May have had a long, backwards-curving crest similar to that of Parasaurolophus
|

|
| Chialingosaurus
|
1959
|
Shaximiao Formation (Late Jurassic, Oxfordian to Kimmeridgian)
|
China
|
Had both large spines and smaller plates, similar to Kentrosaurus
|

|
| Chiayusaurus
|
1953
|
Hasandong Formation?, Xinminbao Group (Early Cretaceous, Barremian to Albian?)
|
China
 South Korea? South Korea?
|
Two species have been named, both from teeth. Those of C. lacustris are apparently indistinguishable from those of Euhelopus[28] or Mamenchisaurus[29]
|
|
| Chilantaisaurus
|
1964
|
Ulansuhai Formation (Late Cretaceous, Turonian)
|
China
|
Had a particularly hooked claw on its first finger
|

|
| Chingkankousaurus
|
1958
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
Known from only a scapula. Possibly a tyrannosauroid[30]
|
|
| Chinshakiangosaurus
|
1992
|
Fengjiahe Formation (Early Jurassic, Hettangian)
|
China
|
Had a U-shaped snout that may have supported fleshy cheeks, an adaptation to bulk feeding
|
|
| Choyrodon
|
2018
|
Khuren Dukh Formation (Early Cretaceous, Albian)
|
Mongolia
|
It had an enlarged nose similar to its contemporary Altirhinus, but it is most likely a separate taxon[31]
|

|
| Chuandongocoelurus
|
1984
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
A tetanuran of uncertain relationships
|

|
| Chuanjiesaurus
|
2000
|
Chuanjie Formation (Middle Jurassic, Bathonian)
|
China
|
One of the most derived mamenchisaurids[32]
|

|
| Chuanqilong
|
2014
|
Jiufotang Formation (Early Cretaceous, Barremian to Aptian)
|
China
|
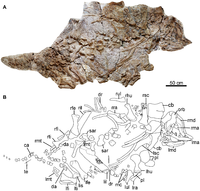
May have been the adult form of the coeval Liaoningosaurus[33]
|
|
| Chungkingosaurus
|
1983
|
Shaximiao Formation (Late Jurassic, Oxfordian)
|
China
|
May have possessed at least six thagomizer spikes. The rearmost pair was mounted horizontally, directed outwards and backwards
|

|
| Chuxiongosaurus
|
2010
|
Lufeng Formation (Early Jurassic, Hettangian to Pliensbachian)
|
China
|
Potentially a synonym of Jingshanosaurus[34]
|
|
| Citipati
|
2001
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Had a distinctive triangular crest. A referred specimen known as the Zamyn Khondt oviraptorid possessed the familiar rectangular domed crest in most depictions of Oviraptor, but likely does not belong to that genus or Citipati[35]
|

|
| Conchoraptor
|
1986
|
Barun Goyot Formation, Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Named for a hypothesized diet of shellfish, but this cannot be confirmed
|

|
| Corythoraptor
|
2017
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Its crest was vertical and rectangular, not unlike that of a cassowary
|

|
| Crichtonpelta
|
2015
|
Sunjiawan Formation (Late Cretaceous, Cenomanian)
|
China
|
Originally named as a second species of Crichtonsaurus
|
|
| Crichtonsaurus
|
2002
|
Sunjiawan Formation (Late Cretaceous, Cenomanian to Turonian)
|
China
|
Sometimes reconstructed with semicircular osteoderms vaguely similar to the plates of stegosaurs
|

|
| Daanosaurus
|
2005
|
Shaximiao Formation (Late Jurassic, Oxfordian to Tithonian)
|
China
|
Its phylogenetic position is uncertain as it is only known from the remains of a juvenile
|
|
| Daliansaurus
|
2017
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Had an enlarged claw on the fourth toe comparable in size to the sickle claw on its second toe
|

|
| Dashanpusaurus
|
2005
|
Shaximiao Formation (Middle Jurassic, Callovian)
|
China
|
One of the basalmost and earliest known macronarians[36]
|
|
| Datai
|
2024
|
Zhoutian Formation (Late Cretaceous, Turonian to Coniacian)
|
China
|
Known from two associated specimens, including their skulls
|

|
| Datanglong
|
2014
|
Xinlong Formation (Early Cretaceous, Barremian to Albian)
|
China
|
Had a uniquely pneumatized ilium similar to megaraptorans
|
|
| Datonglong
|
2016
|
Huiquanpu Formation (Late Cretaceous, Cenomanian to Campanian)
|
China
|
The precise dating of its remains is uncertain
|
|
| Datousaurus
|
1984
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
One of the rarest sauropods of the Shaximiao Formation, known from only two skeletons and a large, deep skull
|

|
| Daurlong
|
2022
|
Longjiang Formation (Early Cretaceous, Aptian)
|
China
|
Preserves remains of an intestinal tract
|

|
| Daxiatitan
|
2008
|
Hekou Group (Early Cretaceous, Barremian)
|
China
|
Large and relatively long-necked
|

|
| Deinocheirus
|
1970
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Had a suite of unique features, most notably a hump supported by elongated neural spines
|

|
| Dilong
|
2004
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Preserves evidence of a coating of simple feathers
|

|
| Dongbeititan
|
2007
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
A theropod tooth has been found encrusted in one of its ribs[37]
|

|
| Dongyangopelta
|
2013
|
Chaochuan Formation (Early Cretaceous to Late Cretaceous, Albian to Cenomanian)
|
China
|
Coexisted with Zhejiangosaurus but could be distinguished based on subtle osteological features[38]
|
|
| Dongyangosaurus
|
2008
|
Jinhua Formation (Late Cretaceous, Turonian to Coniacian)
|
China
|
Its phylogenetic placement is uncertain
|

|
| Dzharaonyx
|
2022
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
One of the oldest known parvicursorines
|
|
| Dzharatitanis
|
2021
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
Originally described as a rebbachisaurid[39] but later reinterpreted as a titanosaur with possible lognkosaurian affinities[40]
|

|
| Elmisaurus
|
1981
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
One of the most complete caenagnathids known
|

|
| Embasaurus
|
1931
|
Neocomian Sands (Early Cretaceous, Berriasian)
|
Kazakhstan
|
Known from only two vertebrae
|

|
| Enigmosaurus
|
1983
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Had a large, backwards-pointing pelvis
|

|
| Eomamenchisaurus
|
2008
|
Zhanghe Formation (Middle Jurassic to Late Jurassic, Aalenian to Oxfordian)
|
China
|
One of the oldest mamenchisaurids
|
|
| Eosinopteryx
|
2013
|
Tiaojishan Formation (Late Jurassic, Oxfordian)
|
China
|
Described as lacking advanced tail feathers and long "hind wings", unlike other paravians, but this may be an artifact of preservation[41]
|

|
| Epidexipteryx
|
2008
|
Haifanggou Formation (Middle Jurassic, Callovian)
|
China
|
Supported four long feathers coming out from an abbreviated tail
|

|
| Equijubus
|
2003
|
Xinminbao Group (Early Cretaceous, Albian)
|
China
|
A grazer that preserves the oldest known evidence of grass-eating[42]
|

|
| Erketu
|
2006
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
May have had the longest neck of any dinosaur relative to its body
|

|
| Erliansaurus
|
2002
|
Iren Dabasu Formation (Late Cretaceous, Cenomanian)
|
China
|
Had long, curved claws on its fingers
|

|
| Erlikosaurus
|
1980
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Preserves the most complete skull known from any therizinosaur
|

|
| Eshanosaurus
|
2001
|
Lufeng Formation (Early Jurassic, Hettangian)
|
China
|
Has been suggested to be the oldest known therizinosaur
|

|
| Euhelopus
|
1956
|
Meng-Yin Formation (Early Cretaceous, Berriasian to Valanginian)
|
China
|
Originally believed to have lived in a marshy environment
|

|
| Euronychodon
|
1991
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
The type species was found in Portugal. The Asian species may represent a form taxon of improperly developed teeth[43]
|
|
| Ferganasaurus
|
2003
|
Balabansai Formation (Middle Jurassic, Callovian)
|
Kyrgyzstan
|
Claimed to have two hand claws, but this has been disputed[44]
|
|
| Ferganocephale
|
2005
|
Balabansai Formation (Middle Jurassic, Callovian)
|
Kyrgyzstan
|
Unusually, its teeth were not serrated
|
|
| Fujianvenator
|
2023
|
Nanyuan Formation (Late Jurassic, Tithonian)
|
China
|
Possessed proportionally long legs which may be an adaptation to wading
|

|
| Fukuiraptor
|
2000
|
Kitadani Formation, Sebayashi Formation? (Early Cretaceous, Barremian to Aptian)
|
Japan
|
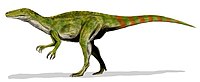
Similarly to Megaraptor, it was originally reconstructed as a dromaeosaur with its hand claw on its foot
|
|
| Fukuisaurus
|
2003
|
Kitadani Formation (Early Cretaceous, Barremian)
|
Japan
|
The elements of its skull are so strongly fused that it was unable to chew[45]
|

|
| Fukuititan
|
2010
|
Kitadani Formation (Early Cretaceous, Barremian to Aptian)
|
Japan
|
The first sauropod named from Japan
|

|
| Fukuivenator
|
2016
|
Kitadani Formation (Early Cretaceous, Barremian to Aptian)
|
Japan
|
Possesses traits of various groups of coelurosaurs, although it may probably be a therizinosaur.[46] May have been a herbivore or omnivore due to its heterodont dentition
|

|
| Fulengia
|
1977
|
Lufeng Formation (Early Jurassic, Hettangian to Toarcian)
|
China
|
May have been a juvenile Lufengosaurus
|
|
| Fushanosaurus
|
2019
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Known from a single femur of immense size
|
|
| Fusuisaurus
|
2006
|
Xinlong Formation (Early Cretaceous, Aptian to Albian)
|
China
|
A referred humerus may support an extremely large size for this taxon[47]
|
|
| Gallimimus
|
1972
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Had a relatively long beak with a rounded tip
|

|
| Gandititan
|
2024
|
Zhoutian Formation (Late Cretaceous, Cenomanian to Turonian)
|
China
|
Possibly a close relative of the Mongolian Abdarainurus[48]
|

|
| Gannansaurus
|
2013
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Its vertebrae were more similar to those of Euhelopus than to other sauropods
|

|
| Ganzhousaurus
|
2013
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Coexisted with at least seven other oviraptorosaurs, which may have niche-partitioned. It was likely primarily herbivorous[49]
|

|
| Garudimimus
|
1981
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Was not as well-adapted to running as later ornithomimosaurs
|

|
| Gasosaurus
|
1985
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
Discovered as a byproduct of construction work
|

|
| Gigantoraptor
|
2007
|
Iren Dabasu Formation (Late Cretaceous, Cenomanian)
|
China
|
The largest known oviraptorosaur, comparable in size to Albertosaurus
|

|
| Gigantspinosaurus
|
1992
|
Shaximiao Formation (Late Jurassic, Oxfordian)
|
China
|
Possessed broad, greatly enlarged shoulder spines
|

|
| Gilmoreosaurus
|
1979
|
Bissekty Formation?, Iren Dabasu Formation, Khodzhakul Formation? (Late Cretaceous, Cenomanian)
|
China
Uzbekistan?
|
Several fossils preserve evidence of cancer-induced tumors[50]
|

|
| Gobihadros
|
2019
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Known from multiple specimens representing different growth stages
|

|
| Gobiraptor
|
2019
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
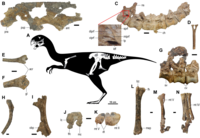
Possessed a deep jaw that may be an adaptation to crushing bivalves or seeds[51]
|
|
| Gobisaurus
|
2001
|
Miaogou Formation (Early Cretaceous, Aptian to Albian)
|
China
|
Had no tail club but already possessed the stiff tail of derived ankylosaurids[52]
|

|
| Gobititan
|
2003
|
Xinminbao Group (Early Cretaceous, Aptian)
|
China
|
Retained the fifth digit of the foot, a basal trait
|
|
| Gobivenator
|
2014
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
The most completely known Cretaceous troodontid
|

|
| Gongbusaurus
|
1983
|
Shaximiao Formation (Late Jurassic, Oxfordian)
|
China
|
Only known from a pair of teeth. May be an ankylosaurian[53]
|
|
| Gongpoquansaurus
|
2014
|
Xinminbao Group (Early Cretaceous, Albian)
|
China
|
Remains originally named as a species of Probactrosaurus
|

|
| Gongxianosaurus
|
1998
|
Ziliujing Formation (Early Jurassic, Toarcian)
|
China
|
The only sauropod with ossified distal tarsals, hinting at its basal position
|
|
| Goyocephale
|
1982
|
Unnamed formation (Late Cretaceous, Campanian)
|
Mongolia
|
Had a sloping head with a flat skull roof
|

|
| Graciliceratops
|
2000
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Possessed a short frill with large fenestrae
|

|
| Graciliraptor
|
2004
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
A close relative of Microraptor with characteristically slender bones
|

|
| Guanlong
|
2006
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Two specimens have been discovered, one on top of the other
|

|
| Halszkaraptor
|
2017
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Originally interpreted as a semiaquatic fish hunter similar to a merganser[54] but this hypothesis has been criticized[55]
|
|
| Hamititan
|
2021
|
Shengjinkou Formation (Early Cretaceous, Aptian)
|
China
|
Known from seven caudal vertebrae and associated elements
|

|
| Haplocheirus
|
2010
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Possessed three long fingers with short claws. Originally described as a basal alvarezsauroid but similarities have been noted with other coelurosaurs[56][57]
|

|
| Harenadraco
|
2024
|
Barun Goyot Formation (Late Cretaceous, Campanian?)
|
Mongolia
|
The first troodontid described from the Barun Goyot Formation[58]
|
|
| Harpymimus
|
1984
|
Khuren Dukh Formation?/Shinekhudag Formation? (Early Cretaceous, Albian)
|
Mongolia
|
Mostly toothless but retains a few teeth in the dentary
|

|
| Haya
|
2011
|
Javkhlant Formation (Late Cretaceous, Santonian to Campanian)
|
Mongolia
|
One specimen preserves a large mass of gastroliths
|

|
| Heishansaurus
|
1953
|
Minhe Formation (Late Cretaceous, Campanian to Maastrichtian)
|
China
|
May be a junior synonym of Pinacosaurus[59]
|
|
| Helioceratops
|
2009
|
Quantou Formation (Early Cretaceous to Late Cretaceous, Albian to Cenomanian)
|
China
|
Had a distinctively short lower jaw
|

|
| Hexing
|
2012
|
Yixian Formation (Early Cretaceous, Valanginian to Barremian)
|
China
|
Three or four teeth are known, but they are not well-preserved
|

|
| Hexinlusaurus
|
2005
|
Shaximiao Formation (Middle Jurassic, Bajocian?)
|
China
|
Originally named as a species of Yandusaurus
|

|
| Heyuannia
|
2002
|
Barun Goyot Formation, Dalangshan Formation (Late Cretaceous, Maastrichtian)
|
China
Mongolia
|
Fossilized pigments in referred eggshells suggest they were blue-green[60]
|

|
| Homalocephale
|
1974
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Has been suggested to be a juvenile Prenocephale on account of its flat head,[61] but this is no longer thought to be the case[62]
|

|
| Huabeisaurus
|
2000
|
Huiquanpu Formation (Late Cretaceous, Cenomanian to Maastrichtian)
|
China
|
May be closely related to Tangvayosaurus[63]
|

|
| Hualianceratops
|
2015
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Had a series of bumps around the edge of the beak
|

|
| Huanansaurus
|
2015
|
Nanxiong Formation (Late Cretaceous, Campanian to Maastrichtian)
|
China
|
Possessed a distinctive short trapezoidal crest
|

|
| Huanghetitan
|
2006
|
Haoling Formation, Hekou Group (Early Cretaceous, Aptian to Albian)
|
China
|
Had ribs estimated to be 3 metres (9.8 ft) long, which supported one of the deepest body cavities of any dinosaur[64]
|

|
| Huangshanlong
|
2014
|
Hongqin Formation (Middle Jurassic to Late Jurassic, Aalenian to Oxfordian)
|
China
|
Known from some bones of the right forelimb
|

|
| Huaxiagnathus
|
2004
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
One of the largest known compsognathids
|

|
| Huaxiazhoulong
|
2024
|
Tangbian Formation (Late Cretaceous, Campanian)
|
China
|
Known from a nearly complete, well-preserved skeleton
|

|
| Huayangosaurus
|
1982
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
Possessed flank osteoderms and a small tail club in addition to plates and spikes
|

|
| Hudiesaurus
|
1997
|
Kalaza Formation (Late Jurassic, Tithonian)
|
China
|
Had a butterfly-shaped process on its vertebra
|

|
| Hulsanpes
|
1982
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Closely related to Halszkaraptor but appears to be more cursorial[65]
|

|
| Hypnovenator
|
2024
|
Ohyamashimo Formation (Early Cretaceous, Albian)
|
Japan
|
The first troodontid named from Japan
|

|
| Ichthyovenator
|
2012
|
Grès supérieurs Formation (Early Cretaceous, Aptian)
|
 Laos Laos
|
One of its sacral vertebrae was greatly reduced, giving the illusion of a break in its sail or of two separate sails
|

|
| Incisivosaurus
|
2002
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Two specimens of different ontogenetic stages are known, both with differing types of feathers[66]
|

|
| Irisosaurus
|
2020
|
Fengjiahe Formation (Early Jurassic, Hettangian)
|
China
|
Closely related to Mussaurus[67]
|

|
| Isanosaurus
|
2000
|
Nam Phong Formation (Uncertain age)
|
 Thailand Thailand
|
Originally thought to be from the Late Triassic but it may have actually come from the Early Jurassic[68] or even the Late Jurassic[69]
|

|
| Ischioceratops
|
2015
|
Wangshi Group (Late Cretaceous, Campanian to Maastrichtian)
|
China
|
Noted for its peculiarly-shaped ischium
|
|
| Isisaurus
|
2003
|
Pab Formation (Late Cretaceous, Maastrichtian)
|
Pakistan
|
The only non-avian dinosaur known from both India and mainland Asia
|

|
| Itemirus
|
1976
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
Originally known from a braincase but abundant new remains were described in 2014[70]
|

|
| Jaculinykus
|
2023
|
Barun Goyot Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Was didactyl, with a large first finger and a reduced second finger
|

|
| Jaxartosaurus
|
1937
|
Dabrazhin Formation (Late Cretaceous, Santonian)
|
Kazakhstan
|
Not known from many remains but they are enough to tell that it was a basal lambeosaurine[71]
|

|
| Jeholosaurus
|
2000
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
One specimen is preserved in a curled up position
|
|
| Jianchangosaurus
|
2013
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Several characters of its teeth and jaws are convergently similar to those of ornithischians[72]
|

|
| Jiangjunosaurus
|
2007
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Had two rows of circular or diamond-shaped plates
|

|
| Jiangshanosaurus
|
2001
|
Jinhua Formation (Late Cretaceous, Turonian to Coniacian)
|
China
|
A potential member of the Euhelopodidae[73]
|

|
| Jiangxisaurus
|
2013
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Overall similar to Heyuannia but with a thinner, frailer mandible
|

|
| Jiangxititan
|
2023
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Although originally described as a titanosaur, a later analysis recovers it as a somphospondylian placed outside of that group[48]
|

|
| Jianianhualong
|
2017
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Possessed a subtriangular tail frond made of asymmetrical feathers, although it was most likely flightless
|

|
| Jinbeisaurus
|
2019
|
Huiquanpu Formation (Late Cretaceous, Cenomanian to Maastrichtian)
|
China
|
A medium-sized tyrannosauroid
|
|
| Jinfengopteryx
|
2005
|
Huajiying Formation (Early Cretaceous, Barremian)
|
China
|
May have been capable of some sort of flight[74]
|

|
| Jingiella
|
2024
|
Dongxing Formation (Late Jurassic, Kimmeridgian?)
|
China
|
Initially named Jingia but that name is already in use by a moth[75]
|

|
| Jingshanosaurus
|
1995
|
Lufeng Formation (Early Jurassic, Hettangian)
|
China
|
One of the latest-surviving non-sauropod sauropodomorphs
|

|
| Jintasaurus
|
2009
|
Xinminbao Group (Early Cretaceous, Albian)
|
China
|
Known from only the rear half of a skull, including a complete braincase
|

|
| Jinyunpelta
|
2018
|
Liangtoutang Formation (Early Cretaceous to Late Cretaceous, Albian to Cenomanian)
|
China
|
The oldest ankylosaurid known to have a tail club
|

|
| Jinzhousaurus
|
2001
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Its holotype is nearly complete, preserved whole on a single slab
|

|
| Jiutaisaurus
|
2006
|
Quantou Formation (Early Cretaceous to Late Cretaceous, Barremian to Cenomanian)
|
China
|
Named based on eighteen vertebrae
|
|
| Kaijiangosaurus
|
1984
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
Potentially synonymous with other medium-sized Shaximiao theropods
|

|
| Kamuysaurus
|
2019
|
Yezo Group (Late Cretaceous, Maastrichtian)
|
Japan
|
Informally referred to as "Mukawaryu" before its formal description
|

|
| Kansaignathus
|
2021
|
Ialovachsk Formation (Late Cretaceous, Santonian)
|
 Tajikistan Tajikistan
|
The first non-avian dinosaur described from Tajikistan
|

|
| Kazaklambia
|
2013
|
Dabrazhin Formation (Late Cretaceous, Santonian)
|
Kazakhstan
|
Morphologically distinct from other Eurasian lambeosaurines[76]
|

|
| Kelmayisaurus
|
1973
|
Tugulu Group (Early Cretaceous, Valanginian to Albian)
|
China
|
One popular book mentions a giant species belonging to this genus,[77] but this referral may be incorrect
|

|
| Kerberosaurus
|
2004
|
Tsagayan Formation (Late Cretaceous, Maastrichtian)
|
Russia
|
Potentially a close relative of Edmontosaurus[78]
|

|
| Khaan
|
2001
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Two morphotypes of chevrons are known, which may be a sexually dimorphic trait[79]
|

|
| Khulsanurus
|
2021
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Contemporary with Parvicursor but can be distinguished by characters of its caudal vertebrae[80]
|
|
| Kileskus
|
2010
|
Itat Formation (Middle Jurassic, Bathonian)
|
Russia
|
Uncertain if it possesses the head crest as seen in other proceratosaurids
|

|
| Kinnareemimus
|
2009
|
Sao Khua Formation (Early Cretaceous, Barremian)
|
Thailand
|
Potentially one of the oldest ornithomimosaurs
|

|
| Kiyacursor
|
2024
|
Ilek Formation (Early Cretaceous, Aptian)
|
Russia
|
Represents a relict population of Jurassic noasaurids
|

|
| Klamelisaurus
|
1993
|
Shishugou Formation (Middle Jurassic, Callovian)
|
China
|
Close relatives included several referred species of Mamenchisaurus[81]
|

|
| Kol
|
2009
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Had a "hyperarctometatarsus" more strongly pinched than other arctometatarsalian taxa. Described as an alvarezsaurid[82] but has been suggested to be related to Avimimus[83]
|
|
| Koreaceratops
|
2011
|
Tando Beds (Early Cretaceous, Albian)
|
South Korea
|
Possessed elongated neural spines on its caudal vertebrae. Its describers suggest that it was used as a swimming organ,[84] but a later study found it to live in a semiarid environment, making this unlikely[85]
|

|
| Koreanosaurus
|
2011
|
Seonso Conglomerate (Late Cretaceous, Campanian)
|
South Korea
|
Had short but powerful forelimbs, suggesting it may have been a quadruped[86]
|

|
| Koshisaurus
|
2015
|
Kitadani Formation (Early Cretaceous, Barremian to Aptian)
|
Japan
|
Distinguished from other hadrosauroids by the presence of an antorbital fossa
|

|
| Kulceratops
|
1995
|
Khodzhakul Formation (Early Cretaceous, Albian)
|
Uzbekistan
|
Only known from fragments of a jaw and teeth
|

|
| Kulindadromeus
|
2014
|
Ukureyskaya Formation (Middle Jurassic, Bathonian)
|
Russia
|
An ornithischian that preserves evidence of filaments, suggesting that protofeathers were basal to Dinosauria as a whole
|

|
| Kundurosaurus
|
2012
|
Udurchukan Formation (Late Cretaceous, Maastrichtian)
|
Russia
|
May be synonymous with Kerberosaurus[87]
|

|
| Kuru
|
2021
|
Barun Goyot Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Had been informally referred to as "Airakoraptor" prior to its formal description
|

|
| Laiyangosaurus
|
2019
|
Wangshi Group (Late Cretaceous, Maastrichtian)
|
China
|
Some specimens referred to this genus actually belong to kritosaurins and lambeosaurines[88]
|

|
| Lanzhousaurus
|
2005
|
Hekou Group (Early Cretaceous, Barremian)
|
China
|
Possessed the largest known teeth of any dinosaur
|

|
| Leshansaurus
|
2009
|
Shaximiao Formation (Late Jurassic, Oxfordian to Kimmeridgian)
|
China
|
Its braincase is nearly identical to that of Piveteausaurus[89]
|

|
| Levnesovia
|
2009
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
One of the smallest known hadrosauroids[44]
|
|
| Liaoceratops
|
2002
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
One specimen was found without a skull roof, possibly displaced by a predator to eat its brain[90]
|

|
| Liaoningosaurus
|
2001
|
Jiufotang Formation, Yixian Formation (Early Cretaceous, Barremian to Aptian)
|
China
|
One specimen has been interpreted as possessing fork-like teeth, sharp claws and stomach contents including fish, which has been claimed to be evidence of a semi-aquatic, turtle-like lifestyle[91]
|

|
| Liaoningotitan
|
2018
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
The second sauropod named from the Yixian Formation
|
|
| Liaoningvenator
|
2017
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Uniquely preserved with the head curving forwards, differing from the classic theropod "death pose" and the sleeping position of other troodontids
|

|
| Limusaurus
|
2009
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Multiple specimens from different growth stages are known. Juveniles possessed teeth which were lost and replaced with a beak as adults, suggesting a change in diet[92]
|

|
| Lingwulong
|
2018
|
Yanan Formation?/Zhiluo Formation? (Middle Jurassic to Late Jurassic, Aalenian to Oxfordian)
|
China
|
The first confirmed diplodocoid from Asia. Originally considered to date from the Early Jurassic, making it the oldest known neosauropod, but this age has been disputed[93][94]
|

|
| Lingyuanosaurus
|
2019
|
Jiufotang Formation?/Yixian Formation? (Early Cretaceous, Valanginian to Aptian)
|
China
|
Possessed a mix of basal and derived therizinosaurian traits
|

|
| Linhenykus
|
2011
|
Bayan Mandahu Formation (Late Cretaceous, Campanian to Maastrichtian)
|
China
|
Completely monodactyl due to lacking the vestigial second and third fingers of other alvarezsaurids
|

|
| Linheraptor
|
2010
|
Bayan Mandahu Formation (Late Cretaceous, Campanian)
|
China
|
Potentially a synonym of Tsaagan[95]
|

|
| Linhevenator
|
2011
|
Bayan Mandahu Formation (Late Cretaceous, Campanian)
|
China
|
Had a greatly enlarged sickle claw, comparable in size to those of dromaeosaurids
|

|
| Lishulong
|
2024
|
Lufeng Formation (Early Jurassic, Sinemurian to Toarcian)
|
China
|
Had the largest skull of any Chinese basal sauropodomorph
|

|
| Liubangosaurus
|
2010
|
Xinlong Formation (Early Cretaceous, Barremian to Aptian)
|
China
|
Described only as a eusauropod[96] but has since been reinterpreted as a somphospondylian[97]
|
|
| Luanchuanraptor
|
2007
|
Qiupa Formation (Late Cretaceous, Maastrichtian)
|
China
|
The first Asian dromaeosaurid found outside the Gobi Desert and northeastern China. May have been closely related to Adasaurus[56]
|

|
| Lufengosaurus
|
1940
|
Lufeng Formation (Early Jurassic, Hettangian to Sinemurian)
|
China
|
The rib of one specimen preserves the oldest known evidence of collagen proteins[98]
|

|
| Luoyanggia
|
2009
|
Haoling Formation (Early Cretaceous, Aptian to Albian)
|
China
|
Originally believed to date from the Late Cretaceous
|
|
| Machairasaurus
|
2010
|
Bayan Mandahu Formation (Late Cretaceous, Campanian)
|
China
|
Its hand claws are elongated and blade-like in side view
|

|
| Mahakala
|
2007
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Possessed basal traits for a dromaeosaurid. May be a close relative of Halszkaraptor[99]
|

|
| Maleevus
|
1987
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Its only purportedly distinguishing trait is also shared with Pinacosaurus[38]
|
|
| Mamenchisaurus
|
1954
|
Penglaizhen Formation, Shaximiao Formation, Shishugou Formation, Suining Formation (Late Jurassic to Early Cretaceous, Oxfordian to Aptian)
|
China
|
Several species have been named, but most may not belong to this genus[81]
|

|
| Mandschurosaurus
|
1930
|
Grès supérieurs Formation?, Yuliangze Formation (Late Cretaceous, Maastrichtian)
|
China
Laos?
|
One of the first non-avian dinosaurs named from Chinese remains
|

|
| Mei
|
2004
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Two specimens are preserved in bird-like sleeping positions[100]
|

|
| Microceratus
|
2008
|
Unnamed formation (Late Cretaceous, Turonian)
|
China
|
Originally named Microceratops, although that genus name is preoccupied by a wasp
|

|
| Microhadrosaurus
|
1979
|
Nanxiong Formation (Late Cretaceous, Campanian to Maastrichtian)
|
China
|
Reportedly an unusually small hadrosaurid
|
|
| Micropachycephalosaurus
|
1978
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
Had the longest name of any known dinosaur
|

|
| Microraptor
|
2000
|
Jiufotang Formation (Early Cretaceous, Aptian)
|
China
|
Known from over three hundred fossils.[101] Several are well-preserved enough to reveal fine details such as feather covering and an iridescent black coloration[102]
|

|
| Migmanychion
|
2023
|
Longjiang Formation (Early Cretaceous, Aptian)
|
China
|
Its hand combines features of multiple groups of coelurosaurs
|

|
| Minimocursor
|
2023
|
Phu Kradung Formation (Late Jurassic, Tithonian)
|
Thailand
|
The first basal neornithischian known from southeastern Asia
|

|
| Minotaurasaurus
|
2009
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
The holotype skull was excavated illegally, which obscured its true provenance until recently
|

|
| Mongolosaurus
|
1933
|
On Gong Formation (Early Cretaceous, Aptian to Albian)
|
China
|
Known from only scant remains but has been confidently assigned to the Somphospondyli in recent years[97]
|

|
| Mongolostegus
|
2018
|
Dzunbain Formation (Early Cretaceous, Aptian to Albian)
|
Mongolia
|
Informally assigned to the genus Wuerhosaurus before its formal description
|

|
| Monkonosaurus
|
1986
|
Loe-ein Formation?/Lura Formation? (Late Jurassic, Oxfordian to Kimmeridgian?/Early Cretaceous, Albian?)
|
China
|
Poorly known
|
|
| Monolophosaurus
|
1993
|
Shishugou Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
Possessed a short, rectangular crest running along the midline of the skull
|

|
| Mononykus
|
1993
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Proposed to have an anteater-like lifestyle, using its unique forearms to break into termite mounds[103]
|

|
| Mosaiceratops
|
2015
|
Xiaguan Formation (Late Cretaceous, Turonian to Campanian)
|
China
|
Combined features of different groups of basal ceratopsians
|

|
| Nankangia
|
2013
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
May have been specialized in soft foods such as leaves and seeds[104]
|

|
| Nanningosaurus
|
2007
|
Unnamed formation (Late Cretaceous, Maastrichtian)
|
China
|
Potentially a basal lambeosaurine
|
|
| Nanshiungosaurus
|
1979
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Originally misidentified as a sauropod on account of its unusual pelvis
|

|
| Nanyangosaurus
|
2000
|
Xiaguan Formation (Late Cretaceous, Turonian to Campanian)
|
China
|
Completely lost the first digit of its hands
|

|
| Napaisaurus
|
2022
|
Xinlong Formation (Early Cretaceous, Aptian to Albian)
|
China
|
May be closely related to contemporary Thai iguanodonts
|
|
| Natovenator
|
2022
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Possessed a streamlined body and a long, toothed snout, convergently similar to several groups of aquatic vertebrates
|

|
| Nebulasaurus
|
2015
|
Zhanghe Formation (Middle Jurassic, Aalenian to Bajocian)
|
China
|
Only known from a single braincase, but it is enough to tell that it was related to Spinophorosaurus
|

|
| Neimongosaurus
|
2001
|
Iren Dabasu Formation (Late Cretaceous, Cenomanian)
|
China
|
Could extend its arms considerably forwards due to the structure of its shoulder joints[105]
|

|
| Nemegtomaia
|
2005
|
Barun Goyot Formation, Nemegt Formation (Late Cretaceous, Campanian to Maastrichtian)
|
Mongolia
|
One specimen preserves traces of damage by skin beetles[106]
|

|
| Nemegtonykus
|
2019
|
Nemegt Formation (Late Cretaceous, Campanian to Maastrichtian)
|
Mongolia
|
The second alvarezsaurid named from the Nemegt Formation
|

|
| Nemegtosaurus
|
1971
|
Nemegt Formation, Subashi Formation? (Late Cretaceous, Maastrichtian)
|
China?
Mongolia
|
Had a long, low skull similar in proportions to those of diplodocoids
|

|
| Ningyuansaurus
|
2012
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Preserves small oval-shaped structures in its stomach region which may be seeds
|
|
| Nipponosaurus
|
1936
|
Yezo Group (Late Cretaceous, Santonian to Campanian)
|
Russia
|
Discovered on the island of Sakhalin, which was owned by Japan in 1936 but later annexed by Russia
|

|
| Oksoko
|
2020
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Its third finger was so greatly reduced that it was functionally didactyl
|

|
| Olorotitan
|
2003
|
Udurchukan Formation (Late Cretaceous, Maastrichtian)
|
Russia
|
Had a broad, hatchet-shaped crest
|

|
| Omeisaurus
|
1939
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
Members of this genus are characterized by extremely elongated necks
|

|
| Ondogurvel
|
2022
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Known from well-preserved remains of the hands and feet
|

|
| Opisthocoelicaudia
|
1977
|
Nemegt Formation (Late Cretaceous, Campanian to Maastrichtian)
|
Mongolia
|
Walked on its metacarpals due to its complete lack of phalanges
|

|
| Oviraptor
|
1924
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Originally mistakenly thought to be an egg-eater
|

|
| Pachysuchus
|
1951
|
Lufeng Formation (Early Jurassic, Sinemurian to Pliensbachian)
|
China
|
Considered a phytosaur from its original naming until a redescription in 2012[107]
|
|
| Panguraptor
|
2014
|
Lufeng Formation (Early Jurassic, Hettangian to Sinemurian)
|
China
|
The first definitive coelophysoid known from Asia
|
|
| Papiliovenator
|
2021
|
Bayan Mandahu Formation (Late Cretaceous, Campanian)
|
China
|
Had a short, subtriangular skull similar to those of Early Cretaceous troodontids
|

|
| Paralitherizinosaurus
|
2022
|
Yezo Group (Late Cretaceous, Campanian)
|
Japan
|
Had stiffened claws that may have been used to pull vegetation to the mouth[108]
|

|
| Parvicursor
|
1996
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Originally believed to represent a diminutive adult dinosaur, although it was recently reinterpreted as a juvenile[109]
|

|
| Pedopenna
|
2005
|
Haifanggou Formation (Middle Jurassic to Late Jurassic, Callovian to Oxfordian)
|
China
|
Known from a single leg with the impressions of long, symmetrical feathers
|

|
| Peishansaurus
|
1953
|
Minhe Formation (Late Cretaceous, Santonian to Campanian)
|
China
|
Has been compared to thyreophorans and marginocephalians, but it is impossible to determine which assignment is correct
|
|
| Penelopognathus
|
2005
|
Bayin-Gobi Formation (Early Cretaceous, Albian)
|
China
|
Named from a single dentary
|
|
| Phaedrolosaurus
|
1973
|
Tugulu Group (Early Cretaceous, Valanginian to Albian)
|
China
|
May have been a dromaeosaurid[110]
|
|
| Philovenator
|
2012
|
Bayan Mandahu Formation (Late Cretaceous, Campanian)
|
China
|
Closely related to the contemporary Linhevenator[100] but likely represents a separate taxon[111]
|

|
| Phuwiangosaurus
|
1994
|
Sao Khua Formation (Early Cretaceous, Valanginian to Hauterivian)
|
Thailand
|
A large member of the Euhelopodidae[97]
|
|
| Phuwiangvenator
|
2019
|
Sao Khua Formation (Early Cretaceous, Barremian)
|
Thailand
|
Combines features of both allosauroids and coelurosaurs[112]
|

|
| Pinacosaurus
|
1933
|
Bayan Mandahu Formation, Djadochta Formation (Late Cretaceous, Campanian)
|
China
Mongolia
|
May have been capable of producing bird-like vocalizations[113]
|

|
| Plesiohadros
|
2014
|
Alagteeg Formation (Late Cretaceous, Campanian)
|
Mongolia
|
The first hadrosauroid known from the Alagteeg Formation
|
|
| Prenocephale
|
1974
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Had a distinctively conical dome
|

|
| Probactrosaurus
|
1966
|
Miaogou Formation (Early Cretaceous, Aptian to Albian)
|
China
|
The closest relative to the Hadrosauromorpha based on the definition of the group[114]
|

|
| Prodeinodon
|
1924
|
Öösh Formation, Xinlong Formation (Early Cretaceous, Barremian to Aptian)
|
China
Mongolia
|
Potentially a carnosaur[115]
|
|
| Protarchaeopteryx
|
1997
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Usually thought to be a basal oviraptorosaur but one study suggests a basal position within Pennaraptora[56]
|

|
| Protoceratops
|
1923
|
Bayan Mandahu Formation, Djadochta Formation (Late Cretaceous, Campanian)
|
China
Mongolia
|
Its remains are so abundant that it has been nicknamed the "sheep of the Cretaceous"
|

|
| Protognathosaurus
|
1991
|
Shaximiao Formation (Middle Jurassic, Bathonian to Callovian)
|
China
|
Originally named Protognathus, but that name is preoccupied by a beetle[116]
|
|
| Psittacosaurus
|
1923
|
Andakhuduk Formation, Bayin-Gobi Formation, Ejinhoro Formation, Ilek Formation, Jiufotang Formation, Khok Kruat Formation?, Öösh Formation, Qingshan Formation, Tugulu Group, Xinminbao Group, Yixian Formation (Early Cretaceous, Barremian to Albian)
|
China
Mongolia
Russia
Thailand?
|
Known from hundreds of specimens, many of them well-preserved. Lived in a broad range
|

|
| Pukyongosaurus
|
2001
|
Hasandong Formation (Early Cretaceous, Aptian to Albian)
|
South Korea
|
One of its caudal vertebrae has bite marks caused by theropod teeth
|

|
| Qianjiangsaurus
|
2024
|
Zhengyang Formation (Late Cretaceous, Cenomanian to Maastrichtian)
|
China
|
This taxon and Nanningosaurus are the only known hadrosauroids from southern China
|

|
| Qianlong
|
2023
|
Ziliujing Formation (Early Jurassic, Sinemurian)
|
China
|
Associated with fossils of leathery eggs, the oldest of their kind in the world
|

|
| Qianzhousaurus
|
2014
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Has been nicknamed "Pinocchio rex" on account of its elongated snout
|

|
| Qiaowanlong
|
2009
|
Xinminbao Group (Early Cretaceous, Aptian)
|
China
|
Originally described as a brachiosaurid[117] but has since been reinterpreted as a euhelopodid[118]
|

|
| Qijianglong
|
2015
|
Suining Formation (Early Cretaceous, Aptian)
|
China
|
Once believed to date from the Late Jurassic
|
|
| Qingxiusaurus
|
2008
|
Unnamed formation (Late Cretaceous, Maastrichtian)
|
China
|
Known from very limited remains
|
|
| Qinlingosaurus
|
1996
|
Hongtuling Formation?/Shanyang Formation? (Late Cretaceous, Maastrichtian)
|
China
|
Potentially a titanosaur given its age, but this cannot be confirmed
|
|
| Qiupalong
|
2011
|
Qiupa Formation (Late Cretaceous, Maastrichtian)
|
China
|
Referred specimens were found in Canada[119] and Russia,[120] making it one of the few Late Cretaceous non-avian dinosaur taxa known from both Asia and Laramidia
|

|
| Qiupanykus
|
2018
|
Qiupa Formation (Late Cretaceous, Maastrichtian)
|
China
|
May have used its robust thumb claws to crack open oviraptorid eggshells[121]
|
|
| Quaesitosaurus
|
1983
|
Barun Goyot Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Potentially a close relative of Nemegtosaurus
|

|
| Ratchasimasaurus
|
2011
|
Khok Kruat Formation (Early Cretaceous, Aptian)
|
Thailand
|
Only known from a single toothless dentary
|

|
| Rhomaleopakhus
|
2021
|
Kalaza Formation (Late Jurassic, Tithonian)
|
China
|
Possessed a robust forelimb that may be a locomotory adaptation
|

|
| Rinchenia
|
1997
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Had a tall, domed crest
|

|
| Ruixinia
|
2022
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Its last few caudal vertebrae were fused into a rod-like structure
|
|
| Ruyangosaurus
|
2009
|
Haoling Formation (Early Cretaceous, Aptian to Albian)
|
China
|
Only known from scant remains but it was one of the largest dinosaurs known from Asia
|

|
| Sahaliyania
|
2008
|
Yuliangze Formation (Late Cretaceous, Maastrichtian)
|
China
|
Possibly a synonym of Amurosaurus[122]
|

|
| Saichania
|
1977
|
Barun Goyot Formation, Nemegt Formation (Late Cretaceous, Campanian to Maastrichtian)
|
Mongolia
|
Possessed complicated nasal passages that may have cooled the air it breathed
|

|
| Sanpasaurus
|
1944
|
Ziliujing Formation (Early Jurassic, Toarcian)
|
China
|
Historically conflated with the remains of an ornithischian
|

|
| Sanxiasaurus
|
2019
|
Xintiangou Formation (Middle Jurassic, Bajocian)
|
China
|
The oldest neornithischian known from Asia
|

|
| Sasayamagnomus
|
2024
|
Ohyamashimo Formation (Early Cretaceous, Albian)
|
Japan
|
At least two individuals are known as indicated by the presence of two right nasal bones among the fossil material
|

|
| Saurolophus
|
1912
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
The type species was found in Canada. The Asian species is distinguished by its larger size and backwards-pointing diagonal crest
|

|
| Sauroplites
|
1953
|
Zhidan Group (Early Cretaceous, Barremian to Aptian)
|
China
|
Preserved lying on its back with parts of its armor in an articulated position
|
|
| Saurornithoides
|
1924
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Its hindlimbs were well-developed even as juveniles, suggesting it needed little to no parental care
|

|
| Scansoriopteryx
|
2002
|
Tiaojishan Formation (Middle Jurassic to Late Jurassic, Callovian to Oxfordian)
|
China
|
Was well-adapted for climbing due to the structure of its hands and feet
|

|
| Segnosaurus
|
1979
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Turonian)
|
Mongolia
|
One of the first known therizinosaurs. Its relationships were originally obscure
|

|
| Serikornis
|
2017
|
Tiaojishan Formation (Late Jurassic, Oxfordian)
|
China
|
Possessed simple, wispy feathers similar to those of a Silkie chicken
|

|
| Shamosaurus
|
1983
|
Dzunbain Formation (Early Cretaceous, Aptian to Albian)
|
Mongolia
|
The osteoderms on its head were not separated into obvious tiles as with later ankylosaurids
|
|
| Shanag
|
2007
|
Öösh Formation (Early Cretaceous, Berriasian to Barremian)
|
Mongolia
|
Shows a mixture of traits of various paravian groups
|

|
| Shantungosaurus
|
1973
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
The largest known hadrosaurid
|

|
| Shanxia
|
1998
|
Huiquanpu Formation (Late Cretaceous, Cenomanian to Campanian)
|
China
|
May be synonymous with Tianzhenosaurus[123] and/or Saichania[38]
|
|
| Shanyangosaurus
|
1996
|
Shanyang Formation (Late Cretaceous, Maastrichtian)
|
China
|
Indeterminate but its hollow bones are a synapomorphy for the Coelurosauria. One study suggests an oviraptorosaurian position[56]
|
|
| Shaochilong
|
2009
|
Miaogou Formation (Early Cretaceous, Aptian to Albian)
|
China
|
Had a relatively short maxilla, suggesting a unique ecological role
|

|
| Shenzhousaurus
|
2003
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Preserves pebbles in its thoracic cavity which may be gastroliths
|

|
| Shidaisaurus
|
2009
|
Chuanjie Formation (Middle Jurassic, Aalenian)
|
China
|
Potentially one of the oldest known allosauroids
|

|
| Shishugounykus
|
2019
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Its manus combines features of both alvarezsaurians and more basal coelurosaurs
|

|
| Shixinggia
|
2005
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
Known from a fair amount of postcranial material
|
|
| Shri
|
2021
|
Barun Goyot Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Before its formal description, it was nicknamed "Ichabodcraniosaurus" because its holotype lacked a skull
|

|
| Shuangmiaosaurus
|
2003
|
Sunjiawan Formation (Early Cretaceous, Albian)
|
China
|
Only known from some parts of a skull
|
|
| Shunosaurus
|
1983
|
Shaximiao Formation (Late Jurassic, Oxfordian)
|
China
|
Possessed a small tail club topped by two short spikes
|

|
| Shuvuuia
|
1998
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Displays several adaptations that may point to a nocturnal, owl-like lifestyle[124]
|

|
| Siamodon
|
2011
|
Khok Kruat Formation (Early Cretaceous, Aptian)
|
Thailand
|
May have been closely related to Probactrosaurus[125]
|

|
| Siamosaurus
|
1986
|
Khok Kruat Formation, Sao Khua Formation (Early Cretaceous, Barremian to Aptian)
|
Thailand
|
Only known from teeth. Some spinosaurid postcrania from the same area may be referrable to this genus[126]
|

|
| Siamotyrannus
|
1996
|
Sao Khua Formation (Early Cretaceous, Berriasian to Barremian)
|
Thailand
|
Has been recovered in a variety of positions within Avetheropoda
|

|
| Siamraptor
|
2019
|
Khok Kruat Formation (Early Cretaceous, Aptian)
|
Thailand
|
Possibly the first carcharodontosaurian known from Southeast Asia
|

|
| Sibirotitan
|
2018
|
Ilek Formation (Early Cretaceous, Barremian)
|
Russia
|
Its sacral ribs are star-shaped in dorsal view
|

|
| Siluosaurus
|
1997
|
Xinminbao Group (Early Cretaceous, Barremian to Albian)
|
China
|
Has been suggested to be an indeterminate member of the Cerapoda
|
|
| Silutitan
|
2021
|
Shengjinkou Formation (Early Cretaceous, Aptian)
|
China
|
Known from six cervical vertebrae associated with a pterosaur jaw
|

|
| Similicaudipteryx
|
2008
|
Jiufotang Formation (Early Cretaceous, Aptian to Albian)
|
China
|
Had a short tail ending with a dagger-shaped pygostyle
|

|
| Sinankylosaurus
|
2020
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
Only known from an ilium. Described as an ankylosaur but a recent study doubts this interpretation[127]
|
|
| Sinocalliopteryx
|
2007
|
Yixian Formation (Early Cretaceous, Barremian to Aptian)
|
China
|
Stomach contents indicate a possible preference for volant prey such as dromaeosaurids and early birds[128]
|

|
| Sinocephale
|
2021
|
Ulansuhai Formation (Late Cretaceous, Turonian)
|
China
|
Originally named as a species of Troodon when that genus was thought to be a pachycephalosaur
|
|
| Sinoceratops
|
2010
|
Wangshi Group (Late Cretaceous, Campanian to Maastrichtian)
|
China
|
Possessed forward-curving hornlets and a series of low knobs on the top of the frill
|

|
| Sinocoelurus
|
1942
|
Kuangyuan Series (Late Jurassic, Oxfordian to Tithonian?)
|
China
|
One study considered it to be a potential plesiosaur[129]
|
|
| Sinornithoides
|
1993
|
Ejinhoro Formation (Early Cretaceous, Aptian to Albian)
|
China
|
Preserved in a roosting position, with its head tucked underneath its left wing
|

|
| Sinornithomimus
|
2003
|
Ulansuhai Formation (Late Cretaceous, Turonian)
|
China
|
Formed age-segregated herds as evidenced by a concentration of juvenile skeletons[130]
|

|
| Sinornithosaurus
|
1999
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
One specimen has grooved teeth, suggesting it was venomous[131]
|

|
| Sinosauropteryx
|
1996
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
The first non-avian dinosaur found with direct evidence of feathers. Analysis of melanosomes suggests it had orange-brown and white countershading with a striped tail and a "bandit mask" around its eyes[132]
|

|
| Sinosaurus
|
1940
|
Lufeng Formation (Early Jurassic, Hettangian to Sinemurian)
|
China
|
Had a pair of midline crests similar to Dilophosaurus
|

|
| Sinotyrannus
|
2009
|
Jiufotang Formation (Early Cretaceous, Aptian)
|
China
|
One of the largest known early tyrannosauroids. Closely related to smaller forms such as Proceratosaurus and Guanlong
|

|
| Sinovenator
|
2002
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Some specimens are preserved three-dimensionally
|

|
| Sinraptor
|
1993
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
May have used its teeth like blades to inflict deep wounds in prey
|

|
| Sinusonasus
|
2004
|
Yixian Formation (Early Cretaceous, Hauterivian)
|
China
|
Had distinctive sinusoid nasal bones
|

|
| Sirindhorna
|
2015
|
Khok Kruat Formation (Early Cretaceous, Aptian)
|
Thailand
|
Its fossils were discovered by corn farmers while digging a reservoir
|

|
| Sonidosaurus
|
2006
|
Iren Dabasu Formation (Late Cretaceous, Turonian to Santonian)
|
China
|
One of the smallest known titanosaurs
|

|
| Stegosaurides
|
1953
|
Xinminbao Group (Early Cretaceous, Hauterivian to Albian)
|
China
|
A thyreophoran of uncertain phylogenetic position
|
|
| Suzhousaurus
|
2007
|
Xinminbao Group (Early Cretaceous, Aptian to Albian)
|
China
|
One of the largest Early Cretaceous therizinosaurs
|

|
| Szechuanosaurus
|
1942
|
Kuangyuan Series (Late Jurassic, Oxfordian to Tithonian?)
|
China
|
Only known from teeth and possibly a very fragmentary skeleton
|

|
| Talarurus
|
1952
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Its tail club has been compared to a wicker basket
|

|
| Tambatitanis
|
2014
|
Ohyamashimo Formation (Early Cretaceous, Albian)
|
Japan
|
Possessed disproportionately large chevrons
|

|
| Tangvayosaurus
|
1999
|
Grès supérieurs Formation (Early Cretaceous, Aptian to Albian)
|
Laos
|
Closely related to Phuwiangosaurus
|

|
| Tanius
|
1929
|
Wangshi Group (Late Cretaceous, Campanian to Maastrichtian)
|
China
|
Today known from only a few bones. More fossils were once present but were not collected
|

|
| Taohelong
|
2013
|
Hekou Group (Early Cretaceous, Albian)
|
China
|
Possessed a sacral shield similar to that of Polacanthus
|
|
| Tarbosaurus
|
1955
|
Nemegt Formation, Subashi Formation (Late Cretaceous, Maastrichtian)
|
China
Mongolia
|
An apex predator that hunted large prey. Very similar to Tyrannosaurus
|

|
| Tarchia
|
1977
|
Barun Goyot Formation, Nemegt Formation (Late Cretaceous, Campanian to Maastrichtian)
|
Mongolia
|
One specimen preserves injuries on its ribs and tail, possibly from a fight with a member of its own kind[133]
|

|
| Tatisaurus
|
1965
|
Lufeng Formation (Early Jurassic, Sinemurian)
|
China
|
Potentially a basal thyreophoran
|
|
| Tengrisaurus
|
2017
|
Murtoi Formation (Early Cretaceous, Valanginian to Hauterivian)
|
Russia
|
Closely related to South American titanosaurs
|
|
| Therizinosaurus
|
1954
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Possessed extremely elongated and stiffened hand claws
|

|
| Tianchisaurus
|
1993
|
Toutunhe Formation (Late Jurassic, Oxfordian to Kimmeridgian)
|
China
|
Its description uses the spellings Tianchisaurus and Tianchiasaurus interchangeably, but the former is correct[134]
|
|
| Tianyulong
|
2009
|
Tiaojishan Formation (Late Jurassic, Oxfordian)
|
China
|
Preserves impressions of long bristles down its back, tail and neck
|

|
| Tianyuraptor
|
2009
|
Yixian Formation (Early Cretaceous, Barremian to Aptian)
|
China
|
Combines features of both northern and southern dromaeosaurids. Had unusual proportions
|

|
| Tianzhenosaurus
|
1998
|
Huiquanpu Formation (Late Cretaceous, Cenomanian to Campanian)
|
China
|
May be synonymous with Saichania[38] but the discovery of the species T. chengi casts doubt on this interpretation
|

|
| Tienshanosaurus
|
1937
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Large but basal for a mamenchisaurid[81]
|

|
| Timurlengia
|
2016
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
Its inner ear was specialized for detecting low-frequency sounds[135]
|

|
| Tochisaurus
|
1991
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Known from only a single metatarsus
|
|
| Tonganosaurus
|
2010
|
Yimen Formation (Early Jurassic, Pliensbachian)
|
China
|
Potentially the oldest known mamenchisaurid
|
|
| Tongtianlong
|
2016
|
Nanxiong Formation (Late Cretaceous, Maastrichtian)
|
China
|
The pose of the holotype suggests it died while trying to free itself from mud
|

|
| Tsaagan
|
2006
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Very similar to Velociraptor but differs in some features of the skull[136]
|

|
| Tsagantegia
|
1993
|
Bayan Shireh Formation (Late Cretaceous, Cenomanian to Santonian)
|
Mongolia
|
Had a long, shovel-shaped snout which may indicate a browsing lifestyle[137]
|

|
| Tsintaosaurus
|
1958
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
Originally mistakenly believed to have possessed a unicorn horn-like crest
|

|
| Tugulusaurus
|
1973
|
Tugulu Group (Early Cretaceous, Barremian to Albian)
|
China
|
Potentially an early, Xiyunykus-grade alvarezsaurian[138]
|
|
| Tuojiangosaurus
|
1977
|
Shaximiao Formation (Late Jurassic, Oxfordian to Kimmeridgian)
|
China
|
Possessed two rows of tall, pointed plates, thickened in the center as if they were modified spikes
|

|
| Turanoceratops
|
1989
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
Had a pair of brow horns like ceratopsids but was likely not a member of that family
|

|
| Tylocephale
|
1974
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Only known from a partial skull but it is enough to tell that it had a remarkably tall dome
|

|
| Tyrannomimus
|
2023
|
Kitadani Formation (Early Cretaceous, Aptian)
|
Japan
|
Its ilium is remarkably similar to that of the supposed tyrannosauroid Aviatyrannis
|

|
| Udanoceratops
|
1992
|
Djadochta Formation (Late Cretaceous, Campanian)
|
Mongolia
|
The largest known leptoceratopsid
|

|
| Ultrasaurus
|
1983
|
Gugyedong Formation (Early Cretaceous, Aptian to Albian)
|
South Korea
|
Described as very large but this may be due to misidentification of a bone
|
|
| Ulughbegsaurus
|
2021
|
Bissekty Formation (Late Cretaceous, Turonian)
|
Uzbekistan
|
A 2022 study suggested this taxon could be a large-bodied dromaeosaurid,[139] although the discovery of a maxilla similar to the holotype supports its classification as a carcharodontosaurian as originally described[140]
|

|
| Urbacodon
|
2007
|
Bissekty Formation, Dzharakuduk Formation, Iren Dabasu Formation (Late Cretaceous, Cenomanian to Turonian)
|
China
Uzbekistan
|
The U. itemirensis holotype preserves a gap separating the eight rear teeth from the rest of its teeth
|
|
| Vayuraptor
|
2019
|
Sao Khua Formation (Early Cretaceous, Barremian)
|
Thailand
|
Potentially ancestral to megaraptorans[141] or an early member of the group[142]
|
|
| Velociraptor
|
1924
|
Bayan Mandahu Formation, Djadochta Formation (Late Cretaceous, Campanian)
|
China
Mongolia
|
One potential specimen preserves quill knobs[143]
|

|
| Wakinosaurus
|
1992
|
Sengoku Formation (Early Cretaceous, Valanginian to Barremian)
|
Japan
|
May be a close relative of Acrocanthosaurus[115]
|
|
| Wannanosaurus
|
1977
|
Xiaoyan Formation (Late Cretaceous, Maastrichtian)
|
China
|
Basal for a pachycephalosaur as indicated by its flat skull with large openings
|

|
| Wuerhosaurus
|
1973
|
Ejinhoro Formation, Tugulu Group (Early Cretaceous, Hauterivian)
|
China
|
One of the last and largest known stegosaurs. Preserved with low rectangular plates but these may be broken
|

|
| Wulagasaurus
|
2008
|
Yuliangze Formation (Late Cretaceous, Maastrichtian)
|
China
|
A rare hadrosaurid known from far less remains than the contemporary Sahaliyania
|

|
| Wulatelong
|
2013
|
Bayan Mandahu Formation (Late Cretaceous, Campanian)
|
China
|
Known from a partial skeleton including some parts of the skull
|

|
| Wulong
|
2020
|
Jiufotang Formation (Early Cretaceous, Aptian)
|
China
|
Analysis of preserved melanosomes suggests it was mostly gray with iridescent wings[144]
|

|
| Xianshanosaurus
|
2009
|
Haoling Formation (Early Cretaceous, Aptian to Albian)
|
China
|
May have been closely related to Daxiatitan[97]
|

|
| Xiaosaurus
|
1983
|
Shaximiao Formation (Middle Jurassic, Bajocian to Callovian)
|
China
|
An ornithischian of uncertain affinities
|
|
| Xiaotingia
|
2011
|
Tiaojishan Formation (Middle Jurassic to Late Jurassic, Bathonian to Oxfordian)
|
China
|
Well-preserved but inconsistent in phylogenetic placement. Some studies suggest a position as an early avialan[145]
|

|
| Xingtianosaurus
|
2019
|
Yixian Formation (Early Cretaceous, Barremian)
|
China
|
Retained the large third finger that was lost in other caudipterids
|

|
| Xingxiulong
|
2017
|
Lufeng Formation (Early Jurassic, Hettangian)
|
China
|
Possessed a robust scapula which increased forelimb mobility for feeding
|

|
| Xinjiangovenator
|
2005
|
Tugulu Group (Early Cretaceous, Valanginian to Albian)
|
China
|
Remains originally identified as Phaedrolosaurus
|

|
| Xinjiangtitan
|
2013
|
Qiketai Formation (Middle Jurassic, Callovian)
|
China
|
Had an extremely long neck
|

|
| Xiongguanlong
|
2009
|
Xinminbao Group, (Early Cretaceous, Aptian)
|
China
|
More robust than other early tyrannosauroids, possibly to support its elongated skull
|

|
| Xixianykus
|
2010
|
Majiacun Formation (Late Cretaceous, Coniacian to Santonian)
|
China
|
One of the smallest known non-avian dinosaurs
|
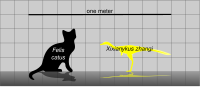
|
| Xixiasaurus
|
2010
|
Majiacun Formation (Late Cretaceous, Coniacian to Campanian)
|
China
|
Distinguished from other troodontids by its possession of exactly twenty-two teeth in each maxilla
|

|
| Xixiposaurus
|
2010
|
Lufeng Formation (Early Jurassic, Hettangian to Toarcian)
|
China
|
Poorly known
|
|
| Xiyunykus
|
2018
|
Tugulu Group (Early Cretaceous, Barremian to Aptian)
|
China
|
Had an unspecialized hand morphology for an alvarezsaur, having three fingers of roughly equal length and construction
|

|
| Xuanhanosaurus
|
1984
|
Shaximiao Formation (Middle Jurassic, Bathonian)
|
China
|
Originally mistakenly believed to have been capable of quadrupedal locomotion
|

|
| Xuanhuaceratops
|
2006
|
Houcheng Formation (Late Jurassic, Tithonian)
|
China
|
Possessed a large premaxillary tooth right behind its beak
|

|
| Xunmenglong
|
2019
|
Huajiying Formation (Early Cretaceous, Hauterivian)
|
China
|
The holotype was originally presented as part of a chimera involving three different animals[146]
|

|
| Xuwulong
|
2011
|
Xinminbao Group (Early Cretaceous, Aptian to Albian)
|
China
|
The tip of its dentary was V-shaped when viewed from the side
|

|
| Yamaceratops
|
2006
|
Javkhlant Formation (Late Cretaceous, Santonian)
|
Mongolia
|
Possessed a short, stubby frill
|

|
| Yamatosaurus
|
2021
|
Kita-Ama Formation (Late Cretaceous, Maastrichtian)
|
Japan
|
Basal yet survived late enough to be contemporaneous with more advanced hadrosaurids
|

|
| Yanbeilong
|
2024
|
Zuoyun Formation (Early Cretaceous, Albian)
|
China
|
One of the youngest known stegosaurs[147]
|

|
| Yandusaurus
|
1979
|
Shaximiao Formation (Middle Jurassic, Bathonian)
|
China
|
Some fossils were destroyed by a composter before they could be studied[148]
|

|
| Yangchuanosaurus
|
1978
|
Shaximiao Formation (Middle Jurassic to Late Jurassic, Bathonian to Tithonian)
|
China
|
The largest theropod known from the Shaximiao Formation
|

|
| Yi
|
2015
|
Tiaojishan Formation (Middle Jurassic to Late Jurassic, Callovian to Oxfordian)
|
China
|
Possessed a "styliform element" jutting out from its wrist that supported a bat-like membranous wing
|

|
| Yimenosaurus
|
1990
|
Fengjiahe Formation (Early Jurassic, Pliensbachian)
|
China
|
Much of its skeleton is known, including the entirety of the skull
|

|
| Yingshanosaurus
|
1994
|
Shaximiao Formation (Late Jurassic, Kimmeridgian)
|
China
|
Possessed greatly enlarged shoulder spines
|

|
| Yinlong
|
2006
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Its skull displays features of ceratopsians, pachycephalosaurs and heterodontosaurids
|

|
| Yixianosaurus
|
2003
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Inconsistent in phylogenetic placement. Had extremely elongated manual elements
|

|
| Yizhousaurus
|
2018
|
Lufeng Formation (Early Jurassic, Sinemurian)
|
China
|
Its skull was very similar to those of sauropods despite being more primitive
|

|
| Yongjinglong
|
2014
|
Hekou Group (Early Cretaceous, Albian)
|
China
|
Possessed an extremely long, broad scapula
|
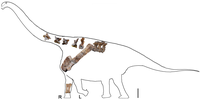
|
| Yuanmousaurus
|
2006
|
Zhanghe Formation (Middle Jurassic, Aalenian to Callovian)
|
China
|
Shares features of its vertebrae with Patagosaurus
|
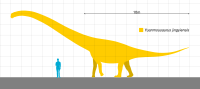
|
| Yuanyanglong
|
2024
|
Miaogou Formation (Early Cretaceous, Aptian to Albian)
|
China
|
The only Early Cretaceous oviraptorosaur known from the Gobi Desert
|

|
| Yueosaurus
|
2012
|
Liangtoutang Formation (Early Cretaceous to Late Cretaceous, Albian to Cenomanian)
|
China
|
Probably closely related to Jeholosaurus[149]
|

|
| Yulong
|
2013
|
Qiupa Formation (Late Cretaceous, Maastrichtian)
|
China
|
Known from multiple specimens, most of which are juveniles
|

|
| Yunganglong
|
2013
|
Zhumapu Formation (Late Cretaceous, Cenomanian)
|
China
|
Discovered 50 kilometres (31 mi) away from a World Heritage Site
|
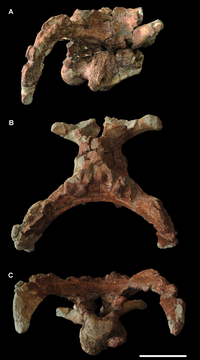
|
| Yunmenglong
|
2013
|
Haoling Formation (Early Cretaceous, Barremian to Albian)
|
China
|
May have been exceptionally large
|
|
| Yunnanosaurus
|
1942
|
Fengjiahe Formation, Lufeng Formation (Early Jurassic, Sinemurian to Pliensbachian)
|
China
|
Its teeth were self-sharpening similar to those of sauropods, likely through convergent evolution[150]
|

|
| Yunyangosaurus
|
2020
|
Xintiangou Formation (Middle Jurassic, Aalenian to Callovian)
|
China
|
Potentially an early megalosauroid
|
|
| Yutyrannus
|
2012
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
The largest known dinosaur to preserve direct evidence of feathers
|

|
| Yuxisaurus
|
2022
|
Fengjiahe Formation (Early Jurassic, Sinemurian to Toarcian)
|
China
|
Had more than one hundred osteoderms
|

|
| Yuzhoulong
|
2022
|
Shaximiao Formation (Middle Jurassic, Bathonian)
|
China
|
One of the oldest known macronarians
|

|
| Zanabazar
|
2009
|
Nemegt Formation (Late Cretaceous, Maastrichtian)
|
Mongolia
|
Originally named as a species of Saurornithoides. Relatively large for a troodontid
|

|
| Zaraapelta
|
2014
|
Barun Goyot Formation (Late Cretaceous, Campanian)
|
Mongolia
|
Had an intricate pattern of osteoderms on its skull
|
|
| Zhanghenglong
|
2014
|
Majiacun Formation (Late Cretaceous, Santonian)
|
China
|
Reconstructed by its describers with a straight, rectangular back, although no complete neural spines are known[151]
|
|
| Zhejiangosaurus
|
2007
|
Chaochuan Formation (Late Cretaceous, Cenomanian)
|
China
|
Has no diagnostic features[38]
|

|
| Zhenyuanlong
|
2015
|
Yixian Formation (Early Cretaceous, Aptian)
|
China
|
Possessed large wings with long feathers, but was most likely flightless
|

|
| Zhongjianosaurus
|
2017
|
Yixian Formation (Early Cretaceous, Barremian to Aptian)
|
China
|
Distinguishable by its characteristically elongated legs. Described as a microraptorian[152] but it has been noted that some features of its skeleton are similar to avialans[41]
|

|
| Zhuchengceratops
|
2010
|
Wangshi Group (Late Cretaceous, Maastrichtian)
|
China
|
Had a particularly deep mandible
|

|
| Zhuchengtitan
|
2017
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
The proportions of its humerus suggest a close relationship with Opisthocoelicaudia[153]
|

|
| Zhuchengtyrannus
|
2011
|
Wangshi Group (Late Cretaceous, Campanian)
|
China
|
Closely related to Tarbosaurus and Tyrannosaurus
|

|
| Zigongosaurus
|
1976
|
Shaximiao Formation (Middle Jurassic to Late Jurassic, Bathonian to Tithonian)
|
China
|
May be a species of Mamenchisaurus[154]
|
|
| Zizhongosaurus
|
1983
|
Ziliujing Formation (Early Jurassic, Toarcian)
|
China
|
Poorly known but was most likely basal for a sauropod
|
|
| Zuolong
|
2010
|
Shishugou Formation (Late Jurassic, Oxfordian)
|
China
|
Known from both cranial and postcranial remains
|

|
| Zuoyunlong
|
2017
|
Zhumapu Formation (Late Cretaceous, Cenomanian)
|
China
|
May have been close to the separation between North American and Asian hadrosauroids[155]
|

|













































.jpg)




























































.png)









































.png)

.jpg)

































































































































.jpg)












.png)









































.jpg)






























_(16411826393).jpg)















-PaleozoologicalMuseumOfChina-May23-08.jpg)






